| Issue |
Knowl. Manag. Aquat. Ecosyst.
Number 425, 2024
Riparian ecology and management
|
|
|---|---|---|
| Article Number | 21 | |
| Number of page(s) | 15 | |
| DOI | https://doi.org/10.1051/kmae/2024019 | |
| Published online | 15 November 2024 | |
Research Paper
Spatial patterns of benthic diatom community structure in the largest northwestern river of Cambodia (Sangker River)
1
Graduate School, National University of Battambang, Battambang City, 021402, Battambang Province, Cambodia
2
Laboratory of Freshwater Ecology (ECOFRESH), Department of Science, Faculty of Science Education, Battambang Teacher Education College (BTEC), Battambang City, 021403, Battambang Province, Cambodia
3
Centre de Recherche sur la Biodiversité et l’Environnement (CRBE), Université de Toulouse, CNRS, IRD, Toulouse INP, Université Toulouse 3–Paul Sabatier (UT3), Toulouse, France
* Corresponding author: This email address is being protected from spambots. You need JavaScript enabled to view it.
Received:
20
August
2024
Accepted:
14
August
2024
Abstract
To date, very little is known about the relative importance of physicochemical factors structuring the benthic diatom community in the flood-pulse tropical ecosystems. This paper aims to investigate the spatial distribution of the benthic diatom community in a poorly studied river catchment of the Tonle Sap Basin in Cambodia. Diatom samples were collected at nineteen (19) monitoring sites from 2018 to 2023 along the Sangker River, and the hierarchical clustering technique was applied to characterize the variation in diatom community structure based on the similarity of diatom genera composition. Seventy-eight (78) diatom genera belonging to 36 families, 18 orders, and 5 classes of diatom flora were recorded. Gomphonema Ehrenberg, Nitzschia Hassall, and Navicula Bory were the most dominant diatom genera in the catchment area. Accordingly, three main diatom assemblages (Ia, Ib, and II) were classified, and 11 indicator genera were identified. Indeed, the identified assemblages reflect the change of diatiom community composition from the upstream assemblage (Ia), middle course (Ib), and floodplain assemblage (II). Dissolved oxygen (DO), pH, conductivity (EC), total phosphorus (TP), and silicon dioxide (SiO2) were the physico-chemical determinants of benthic diatom communities, while physical factors (DO, pH, and nutrients) seemed to be more important than chemical factors. Consequently, any disturbances to the physical condition of the river would drive an abrupt shift in the diatom community’s diversity and structure. These findings pave an essential step towards the establishment of sustainable biomonitoring programs in developing countries such as Cambodia to protect aquatic ecosystems from global change.
Key words: Benthic diatom / spatial distribution / biomonitoring / determinants / Tonle Sap Basin
© S. Chrea et al., Published by EDP Sciences 2024
 This is an Open Access article distributed under the terms of the Creative Commons Attribution License CC-BY-ND (https://creativecommons.org/licenses/by-nd/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. If you remix, transform, or build upon the material, you may not distribute the modified material.
This is an Open Access article distributed under the terms of the Creative Commons Attribution License CC-BY-ND (https://creativecommons.org/licenses/by-nd/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. If you remix, transform, or build upon the material, you may not distribute the modified material.
1 Introduction
The world is suffering nowadays from food and energy crises resulting from ecosystem service degradation (Whiting, 2022; Petrosillo et al., 2023). Ecosystem health monitoring and restoration are among the primary tasks and questions raised by scientific communities across the globe (Cooke et al., 2022). Monitoring freshwater ecosystems is of paramount interest in order to maintain the ecological health of the hydrological systems and to establish the basis for the sustainable management of ecosystem services, which are essential for livelihoods and food security for millions of people in the developing world (Boulton, 1999; Lu et al., 2015; Morin et al., 2016; Pan et al., 2021). However, this requires fundamental knowledge of the biological community of the river ecosystems, e.g., taxonomy, structure and dynamics of the community, environmental determinants (Morin et al., 2016). Diverse biological tools have been recommended to monitor the freshwater ecosystems, particularly for river ecosystem health, e.g., fish, macroinvertebrate, phytoplankton (Karr, 1999; Li, Zheng & Liu, 2010). Among those tools, freshwater diatoms are worldwide recognized as good bio-indicators and are used in many countries to assess the ecological status of running water (Kelly et al., 1998; Wu, 1999; Rott, Pipp & Pfister, 2003; Lobo et al., 2004, 2021; Wang, Stevenson & Metzmeier, 2005).
Diatoms are single-celled algae living in aquatic ecosystems and are among the most diversified protists on earth, with over ten thousand species (Mann & Vanormelingen, 2013). Their presence in all water types, their high diversity, and their specific sensitivity to water pollution make diatoms a relevant tool for bioindication. Diatoms have been extensively used as indicators to assess the ecological quality of river ecosystems over the last few decades, especially across Europe. Many studies have shown the important response of diatoms to pollution driven by human activities, e.g., nutrient enrichment, eutrophication. Diatoms are recommended as bio-indicators of aquatic environments in many countries, and more specifically for most European water types. Over many decades, diatom-based tools (namely diatom indices) have been developed and implemented to assess the status of running water in these countries.
Very few study or documentation of benthic diatom flora in Cambodia have been found. In 1970, the first pioneering study of benthic diatom flora was conducted by Japanese research teams to document the diatom taxa in the Mekong water system, including the Tonle Sap Lake (TSL) and Tonle Sap River (TSR). So far, seventy-seven (77) taxa of diatom were recorded from both lentic and lotic ecosystems (TSL and TSR), and thirty-six (36) benthic diatom taxa were documented for the Mekong River mainstream at that time (Ohno et al., 1972). This exploratory study was a primordial study for further scientific investigation of diatoms in the region. No further research work on benthic diatom flora was found after the first study conducted in 1970 in the TSL or in Cambodia. In 2006, the first biomonitoring program was introduced by the Mekong River Commission (MRC), which aimed to assess the ecological health of the Mekong River system. Thus, a series of data collections on benthic diatoms, zooplankton, littoral macro-invertebrates, and benthic macroinvertebrates was performed, and two hundred six (206) benthic diatom taxa were identified and recorded in the regional database (MRC, 2008). The biomonitoring program continued until 2017 in the four riparian countries, basically along the Mekong mainstream. However, there has been no holistic study of benthic diatoms in Tonle Sap systems (MRC, 2019).
In Thailand, different case studies of diatom community patterns were reported, and diatoms were used as bio-indicators to assess the water quality in Thai river systems (Suphan & Peerapornpisal, 2010; Moonsin et al., 2013; Leelahakriengkai & Kunpradid, 2018). Whereas in northern Laos, investigations of the benthic diatom community within the paddy fields recorded one hundred four (104) diatom taxa, while eleven (11) taxa within the recorded species were unknown (Fujita & Ohtuska, 2005) . At the same time, several new species, e.g., Amphipleura vavilovii sp.nov., were reported from the diatom study in northern Laos (Glushchenko & Kulikovskiy, 2017). In Vietnam, more documentation and scientific literature were found compared to Cambodia and Laos, where benthic diatom-based tools were used to assess the water quality in river systems, e.g., the Saigon River (Luu & Duc, 2018). Many new diatom species have been identified and reported subsequently, e.g., O. japonica, O. subovalis, and O. perjaponica comb. nov. (Kulikovskiy et al., 2015).
Among the Mekong Riparian countries, however, the benthic diatom community in Cambodia was the least studied and investigated after the first exploratory survey by Japanese researchers. Until 2019, diatom research was launched again in the TSL by French research institutions to document the diatom taxa and assess the planktonic diatom community dynamics within the TSL systems (Tudesque et al., 2019, 2021, 2023). So far, fifty-six (56) diatom genera have been recorded from the study (Tudesque et al., 2019). This research provided a good foundation for ecological health assessment of Cambodian freshwater systems; this required a holistic bioecological description of the benthic diatom community in the lake and river systems, however, in order to protect the Cambodian freshwater ecosystem facing rapid global change. The current study aimed to describe the spatial distribution of the benthic diatom community along the Sangker River catchment in the Tonle Sap basin. Specifically, the paper aims to provide the first description of the benthic diatom community structure at the genus level while examining its changes in terms of composition, occurrence, abundance, richness, and diversity along the river course and its response to environmental stressors within the river catchment. This information is an essential step for river ecological health assessment and would pave the foundation for further investigation of the diatom community at the species level as well as enriching the inventory of benthic diatom flora in Cambodia.
2 Materials and methods
2.1 Study area
The largest north-western river of Cambodia, the Sangker River (SR), is known for its cultural, socio-economic, and bio-ecological values, which have sustained the livelihoods of Cambodians for many decades (Campbell et al., 2006). The river connects the north-western upland of Cambodia to the Tonle Sap Lake (TSL), which is known for fish and freshwater bird biodiversity (Fig. 1). The river drains a huge north-western part of the TSL basin with a total catchment area of 6051 km2 (ca. 10% of the TSL basin catchment area). Though the river is critically important for Cambodian daily life, scientific knowledge on the river is somehow scarce. Moreover, rapid socio-economic development within the river catchment, such as urbanization, industrialization, and agricultural intensification, in combination with the recent flooded forest fires in the TSL basin in 2016, has posed an important environmental threat to the river basin ecosystem's health (IUCN Cambodia, 2016; Sourn et al., 2022). Thus, the baseline survey of river health status and the river-based assessment tools must be put in place to restore the river ecosystem services.
The Sangker River basin is known as an agricultural catchment, where the head of the catchment is characterized by a mountainous area and extensive agricultural land (e.g., cassava crops, fruit trees, paddy fields) (Fig. 1). When arriving at the midstream, the catchment is composed of mixed landscapes with dense riparian villages, agricultural land, and industrial areas; the river ends up in the TSL floodplain after a flowing distance of 200 km (Oeurng et al., 2019) (Fig. 1). Geologically, the river is composed of a rocky bottom at the head of the basin, while from the middle to the downstream of the river, the bottom is primarily composed of sediment (muddy river bottoms) (Fig. 1). As part of the TSL basin, the river exhibits the same tropical monsoon climate as the lake, with two distinct seasons: wet (May–Oct) and dry (Apr–Nov).
Recent development within the Sangker River catchment is posing a concern to local authorities, while cascade dam development (e.g., Sek Sak, Kahot, and Taorn dams), deforestation, agricultural intensification, and the rapid growth of urban areas are simultaneously affecting the biodiversity and ecological health of the river (Sourn et al., 2022). Thus, water quality monitoring is essential to protecting the river health and livelihoods of people living in the whole basin. Accordingly, nineteen monitoring sites spatially distributed within the catchment were used for the study, while the majority of sites were located along the Sangker mainstream and a few sites were located in the tributaries (Fig. 1). The selection of the sites was based on the geographical condition and environmental gradient of the river, i.e., mountainous areas, upland, and floodplains, to ensure that the studied sites were representative to assess the bethic diatom community diversity and structure within the catchment.
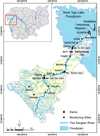 |
Fig. 1 Map of the Sangker River catchment (Tonle Sap Basin, Cambodia), with the geographical location of the nineteen monitoring sites (grey polygones) and the red star indicating the largest city within the catchment area. |
2.2 Benthic diatom data collection, treatment procedure, and taxonomy
The benthic diatoms were collected twice a year in November and in June from 2018 to 2022 within the Sangker River catchment, which aimed primarily to identify the potential locations for long-term environmental monitoring and to develop the diatom-based tools for river ecosystem health assessment. The benthic diatom samples were collected from the available substratum of the river (Fig. 1), in decreasing order of priority: stones (S), macrophyte stems (Sm), macrophyte roots (R), and wood (W). Indeed, to collect the diatom samples, stones and wood were removed from the water and brushed with a toothbrush, while roots and stems were cut and then squeezed. All samples were stored in clean plastic containers (30- 40 ml) and preserved with alcohol (97%). All containers were labeled with the site name, date, and substratum (S, Sm, R, and W) and preserved in a refrigerator (5–7 °C) prior to sample preparation in the labratory.
In the laboratory, all samples were cleaned with boiling hydrogen peroxide (H2O2) and hydrochloric acid (HCl) to remove organic matter. Then, the diatoms were dried onto cover slips and fixed on microscope slides using a high-refractive index resin (Naphrax®, R.I. = 1.7) to obtain permanent slides that can be used for benthic diatom taxonomic identification. Taxonomic identification was done at the genus level by counting the relative abundance of diatoms, i.e., counting 400 valves of diatoms per slide, using the Olympus BX51 microscope (1000× magnification) with oil immersion. The identification of diatom genera was done based on different reference sources, such as diatom identification handbooks from Europe and Asia, published articles, and available benthic diatom studies in the region (Lange-Bertalot, 2001; Krammer, 2002, 2003; Tanaka, 2007, 2014).
2.3 Physico-chemical data collection
Water quality sampling campaigns were conducted where benthic diatoms were collected from November 2018 to June 2022. Twenty-six physico-chemical parameters were measured either in situ or analyzed in the laboratory for each campaign at the nineteen sampling sites. Water temperature (WT), dissolved oxygen (DO), oxygen saturation (OS), electrical conductivity (EC), and pH were measured in-situ using a multi-parameter probe (WTW Multi 3420 Set G), while turbidity (Turb) was measured with a portable turbidity meter (HANNA-HI937030). Other chemical parameters, such as nutrients (total nitrogen (TN), nitrate (NO3–), orthophosphate (PO43–), and various cations and anions), were analyzed in the laboratory in accordance with ISO standard methods according to Chrea et al., 2023. In situ, two 100-ml plastic containers were collected at each site; one container was used to sample raw water for quantifying total nitrogen (TN), and the other was used to store the filtered water (filtered through a 1.2 µm microglass fiber filter). The filters were stored and used to quantify total suspended solids (TSS).
2.4 Statistical analyses
The study aimed to describe the distribution patterns of the benthic diatom community at the genus level. Therefore, the study therefore placed greater emphasis on the composition of genera than on their abundance. The abundance of benthic diatoms was expressed as relative abundance in percentages, while the occurrence was computed as percentages to describe the distribution of the benthic diatom community along the Sangker River. The hierarchical clustering technique was applied to investigate the spatial distribution of the benthic diatom community. Before performing clustering, the benthic diatom table was first averaged from different sampling habitats (stones, macrophyte roots, macrophyte stems, and wood) in each site to reduce the effect of varying sampling efforts between sites, then standardized on the averaged table to reduce the skewness and dominance of certain genera. The standardization was computed using the scale function in package “base” in the R program (R Core Team, 2024), with its default arguments (z = (x − µ)/σ, where z is the standardized or scaled value, x is the original value, µ is the sample mean value, and σ is the sample standard deviation). Next, “Ward.D2” hierarchical clustering was performed based on a standardized benthic diatom table to classify the monitoring sites into different clusters according to their similarity in genera composition (Murtagh & Legendre, 2014). The analysis of similarities (ANOSIM) was then used to test the significant difference in diatom community composition between the identified clusters (Warton et al., 2012). Additionally, genera richness (GR) and diversity index (i.e., Shannon index (SH)) were computed to describe the diversity gradient from the identified clusters, and significant differences (p < 0.05) among clusters were tested using the Kruskal-Wallis test and post-hoc pairwise comparison test.
The indicator genera of each group of sites were then determined using the “indicspecies” package to describe the differences in terms of genara composition within the identified clusters (De Cáceres et al., 2010). Indeed, for a given cluster, the indicator value of the species or genera is the square root of the product of two quantities called A and B, which are predictive value and sensitivity respectively. Quantity A is the probability of the target group of sites given that an individual species has been found and was defined as the mean abundance of the species in the target site group divided by the sum of the mean abundance values over all groups. Quantity B is the average relative abundance of individuals of the species or genera at a site that belongs to the target site group and was determined as the relative frequency of occurrence of the species or genera inside the target site group (De Cáceres et al., 2010). The computation was performed with a permutation of 999 times to ensure the precise combination of diatom genera in each cluster.
To assess the relationship between the benthic diatom community and physicochemical variations, the tri-plot ordination method was applied using canonical correspondence analysis (CCA). CCA is a constraint ordination method that reveals the relationships between community structure, sites, and physicochemical variables (Legendre & Legendre, 2012). Precisely, in the bi-plot ordination map of the CCA model, the importance of physicochemical variables is shown by the length of the vectors, while the correlation between them is exhibited by the angle between the vectors. First step, the CCA model was performed by using all twenty-six contrained physico-chemical factors, then the parsimonious CCA model was computed based on the Monte Carlo permutation test and variance inflation factor (VIF) assessment to improve the performance of the model while reducing the multicollinearity of the constrained factors. Indeed, Monte Carlo permutation tests with 999 permutations were used to test whether the variables significantly (p < 0.05) explained the diatom community structure (Legendre & Legendre, 2012). The “envfit” permutation test with 999 permutations from the “vegan” package was used to examine whether the physicochemical variables significantly influence the variation of the diatom community (Oksanen et al., 2013). Additionally, the “vif.cca” function was used to assess the multicollinearity from all constrained factors (Gross, 2003). Finally, only ten important physicochemical variables were kept for the parsimonious CCA model.
Lastly, to examine the contribution of the two sets of physicochemical factors (i.e., physical vs. chemical factors) in explaining the variation of the benthic diatom community along the river course, variance partitioning was performed to see individual and combined effects from physicochemical factors on the variation of benthic diatom assemblages and was displayed in the form of a Venn Diagram (Legendre & Legendre, 2012; Lai et al., 2022). All statistical analyses were performed in the R program (R Core Team, 2024).
3 Results
3.1 Benthic diatom flora in the Sangker River
Here, the first record of benthic diatom flora from the largest northwestern river in Cambodia, the Sangker River were recorded. So far, seventy-eight (78) genera were recorded from nineteen monitoring sites along the river, belonging to 36 families, 18 orders, and 5 categories of benthic diatoms. Detailed taxonomic descriptions of recorded benthic diatom flora are given in Table S1. In the Sangker River, biraphid diatoms were the most common diatom category found within the catchment, accounting for 67% of diatom genera, followed by centric diatoms (13%), and moniraphid diatoms (10%), while the rest accounted for 10% (Fig. 2a). From the diatom order composition, Naviculales, Cymbellales, and Achnanthales were the most diverse diatoms' orders in the river catchment, accounting for 61% of the recorded total number of genera (Fig. 2b). Accordingly, Naviculaceae was the biggest diatom family, accounting for 10% of genera, followed by Bacillariaceae (8%), Achnanthidiaceae (8%), Cymbellaceae (6%) and Stauroneidaceae (6%), and Staurosiraceae (4%), Diadesmidaceae (4%), Sellaphoraceae (4%), and Stephanodiscaceae (4%) while the rest 42% of families (26 families) contained only one genus, e.g., Ulnariaceae, Amphipleuraceae, Anomoeoneidaceae, Gomphonemataceae, Rhoicospheniaceae, Rhopalodiaceae, Surirellaceae, Thalassiosiraceae, Fragilariaceae, and Tabellariaceae (Fig. 2c).
Indeed, in the Sangker River catchment, approximately 90% of the diatom community's abundance is defined by the thirteen (13) main genera, while the remaining sixty-three (63) genera account for only 10% of their total abundance (Fig. 3a, Tab. S1). More precisely, Gomphonema Ehrenberg (GOMP) was the most abundant genus in the river system, accounting for 19.44% of the relative abundance, followed by Nitzschia Hassall (NITZ) at 17.24%, Navicula Bory (NAVI) at 7.79%, and Achnanthidium Kützing (ACHD) at 7.19% of the relative abundance, respectively (Fig. 3a). Out of the four dominant genera, Sellaphora Mereschowsky (SELL), Cymbella C.Agardh (CYBM), and Eunotia Ehrenberg (EUNO) were among the major diatom genera, accounting for 6.76%, 6.53%, and 5.65% of total abundance, respectively. Other related important genera were Craticula Grunow (CRAT), Melosira C.Agardh (MELO), Encyonopsis Krammer (ENCY), Planothidium Round & L.Bukhtiyarova (PLTD), Brachysira Kützing (BRAC), Ulnaria (Kützing) Compère (ULNA), Fragilaria Lyngbye (FRAG), and Tryblionella W.Smith (TRYB). Each of those genera represented less than 5% of the total abundance, while altogether they accounted for 19.28% of the total abundance. Likewise, GOMP, NITZ, NAVI, ENCY, and FRAG were the most occurring diatom genera along the Sangker river catchment; these genera occurred in all sites and were followed by Luticola D.G.Mann (LUTI) (94.73%), TRYB (94.73%), and CRAT (94.73%), respectively (Fig. 3b).
 |
Fig. 2 Distribution by percentages of benthic diatom genera in the Sangker River catchment (Tonle Sap Basin) according to (a) categories, (b) orders, and (c) families. All families with a number of genera less than 3% were grouped in “Others” and shown in the box next to the pie chart. |
 |
Fig. 3 Abundance and occurrence of benthic diatom community along the Sangker River catchment (Tonle Sap Basin). (a) Relative abundance of diatom genera in percentage (only genera with a relative abundance greater than 1% are shown), (b) Occurrence of diatom genera as a percentage (only diatom genera with a frequency of occurrence greater than 50% are shown). Seventy-eight (78) diatom genera were recorded from the nineteen (19) monitoring sites within the river catchment. |
3.2 Spatial distribution of the benthic diatom community
The nineteen monitoring sites were grouped into three different benthic diatom community clusters based on the similarity of their genus composition (Fig. 4). Two main community clusters were defined at the first split (clusters I and II), reflecting the shift of the river gradient from a lotic to a lentic water system, i.e., the Tonle Sap Lake floodplain. The sub-clusters Ia and Ib revealed the upstream-middle stream's benthic diatom community structure. Accordingly, the analysis of similarity (ANOSIM) revealed a significant difference between the identified clusters (RANOSIM = 0.732, p-value = 0.001). Cluster Ia grouped 8 sites located at the head of the Sangker River basin, while cluster Ib was defined by the majority of sites (6 sites) located in the middle course of the river, and cluster II consisted of 5 sites located in the Tonle Sap floodplain (Fig. 4). A weak gradient of benthic diatom diversity (genera richness and diversity) was observed along the river course. The upstream cluster Ia was characterized by the greatest genera richness (GR = 43.62) and diversity (SH = 2.43), followed by cluster Ib (40.83, 2.36) and downstream cluster II (30.40, 2.24). However, the differences were not found to be significant (Kruskal–Wallis test, p > 0.05).
A total of sixteen benthic diatom indicator genera were identified from the three clusters (Ia, Ib, and II) (Tab. 1, Fig. 5). Consequently, two primary indicator genera, namely Bacillaria J.F.Gmelin (BACI) and Encyonopsis Krammer (ENCP), defined the upstream benthic diatom community (cluster Ia), while the middle course community (Ib) was characterized by two indicator genera, namely Gogorevia Kulikovskiy, Glushchenko, Maltsev & Kociolek (GOGO) and Fistulifera Lange-Bertalot (FITU) (Tab. 1, Fig. 5). In the floodplain, cluster II was characterized by three major indicator genera, namely Frustulia C.Agardh (FRUS), Chamaepinnularia Lange-Bertalot & Krammer (CHAM), and Stenopterobia Brébisson ex Van Heurck (STEN). Additionally, seven indicator genera were identified to characterize the main cluster I (Ia and Ib), i.e., Amphora Ehrenberg ex Kützing (AMPH), CYMB, ACHD, Mayamaea Lange-Bertalot (MAYA), Seminavis D.G.Mann (SMNA), Surirella Turpin (SURI), and Simonsenia Lange-Bertalot (SIMO) (Tab. 1, Fig. 5). Whereas only 2 main indicator genera, i.e., EUNO and Diadesmis Kützing (DIAM), were identified from the combined clusters Ib and II, which are the transitional zones between the river ecosystem and floodplain (Tab. 1, Fig. 5).
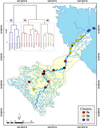 |
Fig. 4 Spatial distribution of the benthic diatom community along the Sangker River catchment (Tonle Sap Basin). Hierarchical clustering (cluster dendrogram) according to the similarity of diatom genera composition (top left). Polygonal dots represent the diatom monitoring sites associated with the identified cluster (Ia & Ib, II). Different colors indicated benthic diatom community assemblages: red (Ia), yellow (Ib), and blue (II), while the black dot represented the dams. |
List of indicator genera of benthic diatom flora by clusters along the Sangker river catchment, Tonle Sap basin (number of permutations = 999).
 |
Fig. 5 Indicator genera of the diatom community are presented by clusters (Ia, Ib, and II) and by combined clusters (Ia and Ib, Ib and II); only the common species within the indicator genera were selected for illustration. |
3.3 Physico-chemical determinants of the benthic diatom community
The CCA model showed an important association between benthic diatom communities and physico-chemical variables (ANOVA test, p = 0.001), while the first two axes of the parsimonious CCA model explained 48.10% of total variances (Fig. 6). Located upstream of the Sangker River, cluster Ia was defined by the genera ENCP and BACI, associated with a high concentration of oxygen (DO and OS) and a high pH (Fig. 6). Cluster Ib, located in the middle stream, was determined by the genera GOGO and FITU, associated with a high concentration of suspended sediment (TSS), nutrients (TP), and silicon dioxide (SiO2) (Fig. 6). Last but not least, cluster II, located in the TSL floodplain, was structured by the genera FRUS, STEN, and CHAM; cluster II was defined by a low concentration of dissolved oxygen, a low pH, and an important concentration of nutrients (TP and NO3), reflecting the downstream condition of the Sangker River dominated by organic pollution from upstream processes (Fig. 6).
 |
Fig. 6 Canonical correspondence analysis (CCA) relating the benthic diatom community (a) to the physicochemical variables (b). Different colors of text on the left plot indicate the indicator genera in each diatom assemblage (yellow text denotes indicator genera from cluster Ia and red and blue from clusters Ib and II, respectively), while the grey texts on the right plot shows the monitoring sites. The blue arrows represent the vectors of physiochemical variables; only significant variables are depicted (Monte Carlo permutation tests with 999 permutations, p-value < 0.05). A detailed description of the benthic diatom genera and Monte Carlo permutation tests are given in Tabs. S1 and S2. |
3.4 Effects of physical and chemical factors on the benthic diatom community
Variation partitioning in the benthic diatom community indicated that physical factors were the predominant factors affecting the variation of diatom community composition along the Sangker River catchment, while the combined effects of both factors (physical and chemical) explained 51% of total variances (Fig. 7). More precisely, physical or chemical factors alone explained 19% and 15% of variances, respectively, while the joint effects of both factors explained 18% of total variances (Fig. 7). Consequently, physical factors seem to be more important than chemical factors in explaining the variation of the diatom community, while the remaining 49% of variances were explained by other factors such as land use/land cover, substratum, altitude, flow velocity, etc.
 |
Fig. 7 Venn diagram of variance partitioning results showing the relative effect of physical and chemical factors alone and in combination with the variation of the benthic diatom community. Numbers represent percentages of variation explained by each factor; all factors were statistically significant (p-value <0.05). |
4 Discussion
4.1 Composition, diversity, and abundance of the benthic diatom community
This study provided the first holistic insight into the benthic diatom community in the largest river in north-western Cambodia, the Sangker River. In total, seventy-eight (78) diatom genera were recorded from the Sangker river, which is higher than the recent record in Tonle Sap Lake (TSL) (i.e., 56 diatom genera) (Tudesque et al., 2019). Naviculales, Cymbellales, and Achnanthales were the most dominant diatom orders in the Sangker river system. Indeed, it was reported in many river systems across Europe, as well as the Mekong River Basin (MRB), the wide distribution of Naviculales and Cymbellales due to their diversities, dispersal ranges, and resistances to pollution (Sala et al., 2008; Rimet & Bouchez, 2012; Larras et al., 2014). The GOMP, NITZ, and NAVI were the most abundant in the river system, accounting for more than 50% of total relative abundances; this was consistent with the previous study reported in TSL (Tudesque et al., 2019). Similarly, from the recent benthic diatom inventory in northern Thailand, researchers recorded fifty-three (53) new species of benthic diatoms, and NAVI, honema Ehrenberg (HONE), and EUNO were the most dominant genera. Besides, other related genera such as Diatomella Greville (DITM), ACHD, Anomoeoneis E.Pfitzer (ANOM), Rhopalodia O.Müller (RHOP), SELL, and CRAT were found among the most abundant diatom genera within the northern river systems of Thailand (Pumas et al., 2018; Suphan & Peerapornpisal, 2010). Whereas in the Saigon River (Vietnam), ACHD, Coscinodiscus Ehrenberg (COSD), and CYMB were found to be among the most dominant genera beside the common genera EUNO, GOMP, NAVI, and NITZ, reflecting the diatom community downstream of the Sangker River cathment (clusters Ib and II), where the quality of habitats is degraded (Pham & Nguyen, 2018; Pham, 2020). Given the fact that GOMP, NAVI, and NITZ are the most rich diatom genera and known as organic pollution-tolerant diatom species, they are usually used for biological monitoring in disturbed tropical river systems (Silva-Benavides, 1996; Hattikudur et al., 2014; Datta et al., 2019).
The gradually decreasing trend of diatom diversity along the Sangker River course reveals the environmental filtering process of benthic diatom community structure (Jamoneau et al., 2018). Consequently, upstream of the river basin (cluster Ia) and middle (cluster Ib) were characterized by high diversity compared to downstream of the river (cluster II). Moreover, benthic diatom flora is very sensitive to disturbance, particularly pollution induced by human activities, as reported in several river systems in Vietnam, Thailand, and Malaysia, with a sharp decline in diversity driven by domestic pollution (Maznah & Mansor, 1999; Thi et al., 2006; Duong et al., 2007; Feio et al., 2009;Masouras et al., 2021). Accordingly, the decreasing trend of diatom diversity was explained by the environmental filtering and organic pollution accumulation processes along the Sangker river course impacted by domestic waste, industrial activities, and agricultural intensification (Heng et al., 2013; Sourn et al., 2022). Moreover, the defined diatom indicator genera from the diatom community are key elements to compute the generic diatom index (GDI), which is one of the most commonly used diatom-based indices for river health assessment (Blanco et al., 2012). Indeed, GDI is a very practical tool to detect organic pollution in river systems, while low values of GDI exhibited low quality of water resulting from the eutrophication effect. Thus, quick and immediate action can be taken to protect ecosystem health, particularly in the data-scarce environment of the Sangker River catchment.
4.2 Relative importance of physico-chemical factors to benthic diatom community structure
Dissolved oxygen (DO), pH, total phosphorus (TP), electrical conductivity (EC), magnesium (Mg2), and silicon dioxide (SiO2) were the most important physico-chemical factors influencing the patterns of the benthic diatom community along the Sangker River catchment. Many previous studies in tropical river systems have demonstrated the relative importance of these physico-chemical factors in explaining the variation of diatom community diversity and structure (Triest et al., 2012; Dalu et al., 2017; Shibabaw et al., 2021). Indeed, oxygen is the predominant factor affecting the distribution of aquatic communities (e.g., fish, invertebrates, phytoplankton, and diatoms) since it relates to the photosynthesis and growth rate of biological communities in the aquatic environment; consequently, more diversity and abundance of diatom communities are expected in the highly oxygenated environment associated with the high level of EC (Tan et al., 2017). Similarly, pH was described as reducing the growth rate and causing physiological stress in the diatom community (Zepernick et al., 2021). Silicon dioxide (SiO2) and magnesium (Mg2) were strongly correlated, while silica is a fundamental element for diatom constitution; thus, the decrease in diatom community was associated with the depletion of SiO2 (Gibson et al., 2000).
The distribution of the benthic diatom community displayed the environmental gradient of the Sangker River. The upstream community (cluster Ia) was defined by the genera BACI and ENCP; these genera have been described to be adapted to marine or brackish water (i.e., BACI) with a high level of EC and nutrients (Spaulding et al., 2021), which is in contrast to the upstream conditions of the Sangker River. The possible explanation for this observed pattern can be associated with the variability of bioecological responses within the species among the genera. For instance, the genus ENCP was described as containing more than 28 species, and several species were reported to be suited to an oligotrophic environment with a high level of DO and low nutrients, e.g., Encyonopsis carraensis sp. nov. (Kennedy et al., 2019). Besides, a recent study from Dongting Lake in China reported the new species of BACI in the freshwater ecosystem, i.e., Bacillaria, B. sinensis sp. nov., and B. dongtingensis sp. nov. (Long et al., 2021). More importantly, cluster Ia was defined by a high concentration of DO and a high level of pH associated with a high concentration of SiO2, revealing the pristine condition at the head of the Sangker River that is suitable to accommodate a great diversity of benthic diatom communities (Sourn et al., 2022).
The middle stream diatom community (cluster Ib) was described by the genera GOGO and FITU; these genera are usually found in eutrophic environments and were described to be resistant to organic pollution (Lange-Bertalot et al., 2018;Spaulding et al., 2021). Consequently, the middle part of the Sangker River was shown to be impacted by industrial activities and agricultural and domestic pollution as a result of socio-economic activities from the riparian villages (Solak et al., 2020). In the TSL floodplain, the diatom community (cluster II) was described by FRUS and STEN. These genera are resistant to a hypoxic environment with a low level of DO (e.g., DO < 2 mg L−1) and a high nutrient concentration. This observed pattern was consistent with the recent benthic diatom study in Tonle Sap Lake and the bioecological description from the USA diatom database (Tudesque et al., 2019; Spaulding et al., 2021). Indeed, FRUS, CHAM, and STEN were known as bio-indicators for siltation in river systems and were frequently observed frequently in acidic environments, as reported in a previous study in French Guiana (Sylvestre et al., 2004; Bey & Ector, 2013; Ríos-Pulgarín et al., 2016).
The current study shows that the benthic diatom community is very sensitive to environmental changes, particularly the impact of physical factors induced by human activities. Changes in physical factors such as DO, pH, EC, and TSS therefore contributed significantly to the variation of the benthic diatom community diversity and structure in the Sangker River system. Indeed, these physical factors have strong effects on diatom community growth, while benthic diatom would colony productively in a less harmful, less turbid, and highly oxygenated environment, as reported in many studies from Europe and Africa (Finkel et al., 2009; Verleyen et al., 2009; Shibabaw et al., 2021). Thus, any disturbance to the physical conditions of the river system would significantly change the diatom community composition, diversity, occurrence, and abundance (Soininen et al., 2014; Shibabaw et al., 2021).
In addition, under the tropical monsoon climate, a major seasonal change in benthic diatom community diversity and composition was expected within the Sangker river catchment, given the fact that environmental conditions change significantly between wet and dry seasons. Consequently, high diversity was expected in the wet season compared to the dry season due to the great species turnover mechanism and habitat diversity in the wet season as reported from the recent case studies in TSL in Cambodia, Yellow River Delta and Pearl River Delta in China (Huang et al., 2004; Ohtaka et al., 2010; Tudesque et al., 2019; Liu et al., 2022). Recently, Yoshikawa et al. (2024) revealed important spatial and seasonal variations of benthic diatom abundance, i.e., Aulacoseira and Suriella, across the Tonle Sap Lake. High abundance was observed in the high water season compared to the low water period, given the fact that the intrusion of river water in the lake induced nutrient enrichment, which are the main drivers of diatom community composition, diversity, and biomass. More evidence on the relevance of benthic diatoms for ecosystem health assesment has been proven; this should be a foundation for future investigations of benthic diatom flora and establishment of biomonitoring metrics for Cambodian freshwater ecosystems (Thi et al., 2006; Wu et al., 2012;Venkatachalapathy & Karthikeyan, 2015). Therefore, detailed bioecological descriptions of Cambodian diatom flora at species level are highly recommended for future studies in order to generate a comprehensive catalogue of diatom flora that is fundamental for long-term environmental monitoring in Cambodia and the Mekong region.
Supplementary Material
Tab. S1. Inventory list of benthic diatom flora in the Sangker River catchment (Tonle Sap basin, Cambodia).
Tab. S2. Environmental determinants of benthic diatom community structure along the Sangker River catchment (environmental fitting model, 999 permutations). Underlined determinants denoted physical factors, while italic ones denoted chemical factors.
Access Supplementary MaterialAcknowledgements
The authors are grateful to the JEAI-AGROECO Project “Preserving ecosystem function and productivity of the Tonle Sap system facing global change” for providing the funds to support the Laboratory of Freshwater Ecology (ECOFRESH) on the biomonitoring of the Sangker River catchment. Great thanks to Eléanor France for English correction in our revised manuscript and thanks to the PHC Tonle Sap Project No. 51509WL for researcher mobility support. Thanks to the Centre de Recherche sur la Biodiversité et l'Environnement (CRBE), Université de Toulouse, CNRS, IRD, Toulouse INP, Université Toulouse 3–Paul Sabatier (UT3), Toulouse, France. The CRBE was supported by ‘Investissement d’Avenir' grants (CEBA, ref. ANR-10-LABX-0025; TULIP, ref. ANR-10-LABX-41).
References
- Bey M-Y, Ector L. 2013. Atlas des diatomées des cours d’eau de la région Rhône-Alpes. Tome 1 Centriques, Monoraphidées. Tome 2 Araphidées, Brachyraphidées. Tome 3 Naviculacées: Naviculoidées. Tome 4 Naviculacées: Naviculoidées. Tome 5 Naviculacées : Cymbelloidées, Gomphonématoid. Direction régionale de l’Environnement, de l’Aménagement et du Logement Rhône-Alpes, Lyon, pp. 1182. [Google Scholar]
- Blanco S, Cejudo-Figueiras C, Tudesque L, Bécares E, Hoffmann L, Ector L. 2012. Are diatom diversity indices reliable monitoring metrics? Hydrobiologia 695: 199–206. [CrossRef] [Google Scholar]
- Boulton AJ. 1999. An overview of river health assessment: philosophies, practice, problems and prognosis. Freshw Biol 41: 469–479. [CrossRef] [Google Scholar]
- De Cáceres M, Legendre P, Moretti M. 2010. Improving indicator species analysis by combining groups of sites. Oikos 119: 1674–1684. [CrossRef] [Google Scholar]
- Campbell IC, Poole C, Giesen W, Valbo-Jorgensen J. 2006. Species diversity and ecology of Tonle Sap Great Lake, Cambodia. Aquatic Sci 68: 355–373 [CrossRef] [Google Scholar]
- Chrea S, Tudesque L, Chea R. 2023. Comparative assessment of water quality classification techniques in the largest north-western river of Cambodia (Sangker River-Tonle Sap Basin). Ecolog Indic 154: 110759. [CrossRef] [Google Scholar]
- Cooke SJ, Frempong-Manso A, Piczak ML, Karathanou E, Clavijo C, Ajagbe SO, et al. 2022. A freshwater perspective on the United Nations decade for ecosystem restoration. Conserv Sci Practice 4: 1–12. [Google Scholar]
- Dalu T, Wasserman RJ, Magoro ML, Mwedzi T, Froneman PW, Weyl OLF. 2017. Variation partitioning of benthic diatom community matrices: effects of multiple variables on benthic diatom communities in an Austral temperate river system. Sci Total Environ 601–602: 73–82. [Google Scholar]
- Datta A, Marella TK, Tiwari A, Wani SP. 2019. The Diatoms: From Eutrophic Indicators to Mitigators BT - Application of Microalgae in Wastewater Treatment: Volume 1: Domestic and Industrial Wastewater Treatment. In: Application of Microalgae in Wastewater Treatment. (Eds S.K. Gupta & F. Bux), pp. 19–40. [Google Scholar]
- Datta A, Marella TK, Tiwari A, Wani SP. 2019. The Diatoms: From Eutrophic Indicators to Mitigators. 19–40. https://doi.org/10.1007/978-3-030-13913-1 [Google Scholar]
- Duong TT, Feurtet-Mazel A, Coste M, Dang DK, Boudou A. 2007. Dynamics of diatom colonization process in some rivers influenced by urban pollution (Hanoi, Vietnam). Ecolog Indic 7: 839–851. [CrossRef] [Google Scholar]
- Feio MJ, Almeida SFP, Craveiro SC, Calado AJ. 2009. A comparison between biotic indices and predictive models in stream water quality assessment based on benthic diatom communities. Ecolog Indic 9: 497–507. [CrossRef] [Google Scholar]
- Finkel ZV, Vaillancourt CJ, Irwin AJ, Reavie ED, Smol JP. 2009. Environmental control of diatom community size structure varies across aquatic ecosystems. Proc Royal Soc B: Biol Sci 276: 1627–1634. [CrossRef] [PubMed] [Google Scholar]
- Fujita Y, Ohtuska T. 2005. Diatoms from paddy fields in northern Laos. Diatom 21: 71–89 [Google Scholar]
- Gibson CE, Wang G, Foy RH. 2000. Silica and diatom growth in Lough Neagh: the importance of internal recycling. Freshw Biol 45: 285–293. [CrossRef] [Google Scholar]
- Glushchenko AM, Kulikovskiy MS. 2017. Amphipleura vavilovii: A New Diatom Species of the Family Amphipleuraceae from Laos. Inland Water Biology 10: 17–21. [Google Scholar]
- Gomà J, Rimet F, Cambra J, Hoffmann L, Ector L. 2005. Diatom communities and water quality assessment in Mountain Rivers of the upper Segre basin (La Cerdanya, Oriental Pyrenees). Hydrobiologia 551: 209–225. [CrossRef] [Google Scholar]
- Gross J. 2003. Variance inflation factors. R News 3: 13–15 [Google Scholar]
- Hattikudur RS, Alakananda B, Ramachandra TV. 2014. Diatoms as bioindicators. LAKE 2014: Conference on Conservation and Sustainable Management of Wetland Ecosystems in Western Ghats, 13–15 November, 2014: 2012–2015. [Google Scholar]
- Heng C, Doch S, Diepart J-C. 2013. Toward measuring the vulnerability of agricultural production to flood: insight from sangkae river catchment, Battambang Province, Cambodia. Int J Environ Rural Dev 4: 89–97. [Google Scholar]
- Huang L, Jian W, Song X, Huang X, Liu S, Qian P, et al. 2004. Species diversity and distribution for phytoplankton of the Pearl River estuary during rainy and dry seasons. Mar Pollut Bull 49: 588–596. [CrossRef] [PubMed] [Google Scholar]
- IUCN Cambodia. 2016. Flooded forest fires: a major threat to the Tonle Sap. News and Events, Phnom Penh, Cambodia. [Google Scholar]
- Jamoneau A, Passy SI, Soininen J, Leboucher T, Tison-Rosebery J. 2018. Beta diversity of diatom species and ecological guilds: response to environmental and spatial mechanisms along the stream watercourse. Freshw Biol 63: 62–73. [CrossRef] [Google Scholar]
- Karr J. 1999. Defining and measuring river health. Freshw Biol 41: 221–234 [CrossRef] [Google Scholar]
- Kelly MG, Cazaubon A, Coring E, Uomo AD, Ector L, Goldsmith B et al. 1998. Recoomendations for the routine sampling of diatoms for water assessment in Europe. 215–224 [Google Scholar]
- Kennedy B, Buckley Y, Allott N. 2019. Taxonomy, ecology and analysis of type material of some small Encyonopsis with description of new species in Ireland. Phytotaxa 395: 89–128. [CrossRef] [Google Scholar]
- Krammer K. 2002. Cymbell. In: Diatoms of Europe 3. (Ed. H. Lange-Bertalot), p. 584. A.R.G. Gantner Verlag K.G., Ruggell. [Google Scholar]
- Krammer K. 2003. Cymbopleura, Delicata, Navicymbula, Gomphocymbellopsis, Afrocymbella. In: Diatoms of Europe 4. (Ed. H. Lange-Bertalot), p. 530. A.R.G. Gantner Verlag K.G., Ruggell. [Google Scholar]
- Kulikovskiy M, Glushchenko A, Kociolek Jo.P. 2015. The diatom genus Oricymba in Vietnam and Laos with description of one new species, and a consideration of its systematic placement. Phytotaxa 227: 120–134. [CrossRef] [Google Scholar]
- Lai J, Zou Y, Zhang J, Peres-Neto PR. 2022. Generalizing hierarchical and variation partitioning in multiple regression and canonical analyses using the rdacca.hp R package. Methods Ecol Evolut 13: 782–788. [CrossRef] [Google Scholar]
- Lange-Bertalot H. 2001. Navicula sensu stricto, 10 genera separated from Navicula sensu lato, Frustulia. In: Diatoms of Europe: Diatoms of the European inland waters and comparable habitats. Germany: A.R.G. Gantner Verlag KG. [Google Scholar]
- Lange-Bertalot H, Hofmann G, Werum M, Cantonati M. 2017. Freshwater Benthic Diatoms of Central Europe: Over 800 common Species Used in Ecological Assessment, Koeltz Botanical, 901 pp. [Google Scholar]
- Larras F, Keck F, Montuelle B, Rimet F, Bouchez A. 2014. Linking diatom sensitivity to herbicides to phylogeny: a step forward for biomonitoring? Environ Sci Technol 48: 1921–1930. [CrossRef] [PubMed] [Google Scholar]
- Leelahakriengkai P, Kunpradid T. 2018. Benthic diatoms in the Ping river and its tributaries in Mae Taeng district, Chiang Mai province, Thailand. Biotropia 9: 181–192. [Google Scholar]
- Legendre P, Legendre L. 2012. Numerical Ecology. Third Engl. Elsevier. [Google Scholar]
- Li L, Zheng B, Liu L. 2010. Biomonitoring and bioindicators used for river ecosystems: definitions, approaches and trends. Proc Environ Sci 2: 1510–1524. [CrossRef] [Google Scholar]
- Liu X, Han J, Li Y, Zhu B, Li Y, Pan K. 2022. The structure of the seasonal benthic diatom community and its relationship with environmental factors in the Yellow River Delta. Front Marine Sci 9: 1–10. [Google Scholar]
- Lobo EA, Bes D, Tudesque L, Ector L. 2004. Water quality assessment of the Pardinho River, RS, Brazil, using epilithic diatom assemblages and faecal coliforms as biological indicators. Vie et Milieu 54: 115–125. [Google Scholar]
- Long J, Williams DM, Liu B, Zhou Y. 2021. Two new freshwater species of Bacillaria (Bacillariophyta) from Dongting Lake, China. Natural Science 20: 1–24. [Google Scholar]
- Lu Y, Wang R, Zhang Y, Su H, Wang P, Jenkins A, et al. 2015. Ecosystem health towards sustainability. Ecosyst Health Sustain 1: 1–15. [Google Scholar]
- Luu PT, Duc NT. 2018. Using benthic diatoms as bio-indicators of water quality of the Saigon River, Vietnam. Environment 54: 106–111. [Google Scholar]
- Mann DG, Vanormelingen P. 2013. An Inordinate Fondness? The Number, Distributions, and Origins of Diatom Species. J Eukaryot Microbiol 414–420. [CrossRef] [PubMed] [Google Scholar]
- Masouras A, Karaouzas I, Dimitriou E, Tsirtsis G, Smeti E. 2021. Benthic diatoms in river biomonitoring-present and future perspectives within the water framework directive. Water (Switzerland) 13. https://doi.org/10.3390/w13040478 [Google Scholar]
- Maznah W, & Mansor M. 1999. Benthic diatoms in the Pinang River (Malaysia) and its tributaries with emphasis on species diversity and water quality. Int J Algae 1: 103–118. [CrossRef] [Google Scholar]
- Moonsin P, Peerapornpisal Y, Rott E, Pipp E, Pimmongkol A. 2013. Diversity of benthic diatoms and water quality of the Mekong River Passing. Res J Environ Earth Sci 5: 58–66. [Google Scholar]
- Morin S, Gómez N, Tornés E, Licursi M, Rosebery J. 2016. Benthic diatom monitoring and assessment of freshwater environments: standard methods and future challenges. Aquatic Biofilms 6: 111–124. [Google Scholar]
- MRC. 2008. Biomonitoring of the lower Mekong River and selected tributaries, 2004–2007. Mekong River Commission, Vientiane. [Google Scholar]
- MRC. 2019. Report on the 2017 biomonitoring survey of the Lower Mekong River and selected tributaries. MRC Technical Paper, The Mekong River Commission/Vientiane/Lao PDR, 102. [Google Scholar]
- Murtagh F, Legendre P. 2014. Ward s hierarchical agglomerative clustering method: which algorithms implement ward ' s criterion? J Classific 31: 274–295. [CrossRef] [Google Scholar]
- Oeurng CA, Cochrane T, Chung S, Kondolf GM, Piman T, Arias EM. 2019. Assessing climate change impacts on river flows in the Tonle Sap Lake Basin, Cambodia. Water (Switzerland) 11: 1–27. [Google Scholar]
- Ohno M, Fukushima H, Ko-Bayashi T. 1972. Diatom Flora of the Mekong Water System, Cambodia. Natural Science 20: 1–24. [Google Scholar]
- Ohno M, Fukushima H, Kobayashi T. 1972. Diatom Flora of the Mekong Water System, Cambodia. Natural Science 20. 1–24. [Google Scholar]
- Ohtaka A, Watanabe R, Im S, Chhay R, Tsukawaki S. 2010. Spatial and seasonal changes of net plankton and zoobenthos in Lake Tonle Sap, Cambodia. Limnology 11: 85–94. [CrossRef] [Google Scholar]
- Oksanen J, Blanchet F.G., Kindt R, Legendre P, Minchin P.R., O’hara R.B, et al. 2013. vegan: Community Ecology Package. Software 2, 1–295 [Google Scholar]
- Pan Z, He J, Liu D, Wang J, Guo X. 2021. Ecosystem health assessment based on ecological integrity and ecosystem services demand in the Middle Reaches of the Yangtze River Economic Belt, China. Sci Total Environ 774: 144837. [CrossRef] [Google Scholar]
- Petrosillo I, Valente D, Scavuzzo CM, Selvan T. 2023. Editorial: Land degradation pattern and ecosystem services. Front Environ Sci 11: 1–4. [CrossRef] [Google Scholar]
- Pham TL. 2020. Using benthic diatoms as a bioindicator to assess Rural-Urban River conditions in tropical area: a case study in the Sai Gon River, Vietnam. Pollution 6: 387–398. [Google Scholar]
- Pham TL, Nguyen TD. 2018. Using benthic diatoms as bio-indicators of water quality of the Saigon River, Vietnam. Can Tho Univ J Sci 54: 106. [Google Scholar]
- Pumas C, Pruetiworanan S, PeeraPornPisal Y. 2018. Diatom diversity in some hot springs of Northern Thailand. Botanica 24: 69–86. [Google Scholar]
- R Core Team. 2024. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. [Google Scholar]
- Rimet F, Bouchez A. 2012. Life-forms, cell-sizes and ecological guilds of diatoms in European rivers. Knowl Manag Aquat Ecosyst 1–12. [CrossRef] [EDP Sciences] [Google Scholar]
- Ríos-Pulgarín MI, Gil-Guarín IC, Barletta M, Mancera-Rodríguez NJ. 2016. Effects of the hydrological cycle on the phycoperiphyton assemblage in an andean foothill stream in Colombia. J Limnol 75: 121–136. [Google Scholar]
- Rott E, Pipp E, Pfister P. 2003. Diatom methods developed for river quality assess- ment in Austria and a cross-check against numerical trophic indication methods used in Europe. Algolog Stud 110: 70176. [Google Scholar]
- Sala SE, Ramírez JJ, Plata Y. 2008. Diatoms from lentic and lotic systems in antioquia, chocó and santander departments in Colombia. Rev Biol Trop 56: 1159–1178. [PubMed] [Google Scholar]
- Shibabaw T, Beyene A, Awoke A, Tirfie M, Azage M, Triest L. 2021. Diatom community structure in relation to environmental factors in human influenced rivers and streams in tropical Africa. PLOS ONE 16: e 0246043. [Google Scholar]
- Silva-Benavides A-M. 1996. The use of water chemistry and benthic diatom communities for qualification of a polluted tropical river in Costa Rica. Rev Biol Trop 44: 395–416. [PubMed] [Google Scholar]
- Soininen J, Paavola R, Muotka T. 2014. in boreal streams: benthic diatom communities structure community in relation to environmental and spatial gradients. Ecography 27: 330–342. [Google Scholar]
- Solak CN, Peszek Ł, Yilmaz E, Ergül HA, Kayal M, Ekmekçi F, et al. 2020. Use of Diatoms in Monitoring the Sakarya River Basin. Turkey Water 12: 703. [Google Scholar]
- Sourn T, Pok S, Chou P, Nut N, Theng D, Prasad PVV. 2022. Assessment of land use and land cover changes on soil erosion using remote sensing, GIS and RUSLE model: a case study of Battambang Province, Cambodia. Sustainability 14: 4066. [CrossRef] [Google Scholar]
- Spaulding SA, Potapova MG, Bishop IW, Lee SS, Gasperak TS, Jovanoska E, et al. 2021. Diatoms.org: supporting taxonomists, connecting communities. Diatom Res 36: 291–304. [CrossRef] [Google Scholar]
- Suphan S, Peerapornpisal Y. 2010. Fifty three new record species of benthic diatoms from mekong river and its tributaries in Thailand. Chiang Mai J Sci 37: 326–343. [Google Scholar]
- Sylvestre F, Guiral D, Debenary J. 2004. Modern diatom distribution in mangrove swamps from the Kaw Estuary (French Guiana). Marine Geology 208: 281–293. [Google Scholar]
- Tan X, Zhang Q, Burford MA, Sheldon F, Bunn SE. 2017. Benthic diatom based indices for water quality assessment in two subtropical streams. Front Microbiol 8, 1–10. [PubMed] [Google Scholar]
- Tanaka H. 2014. Atlas of freshwater fossil diatoms in Japan, including related recent taxa. Tokyo, Japan, Uchida Rokakuho Publishing Co., Ltd. [Google Scholar]
- Tanaka H. 2007. Taxonomic Studies of the Genera Cyclotella (Kuzing) Brebisson, Discotella Houk et Klee and Puncticulata Hakansson in the Family Sephanodiscaceae Glezer et Makarova (Bacillariophyta) in Japan. 5–54 [Google Scholar]
- Thi TD, Coste M, Feurtet-Mazel A, Dinh KD, Gold C, Young SP, et al. 2006. Impact of urban pollution from the Hanoi area on benthic diatom communities collected from the Red, Nhue and Tolich rivers (Vietnam). Hydrobiologia 563: 201–216. [CrossRef] [Google Scholar]
- Triest L, Lung'ayia H, Ndiritu G, Beyene A. 2012. Epilithic diatoms as indicators in tropical African rivers (Lake Victoria catchment). Hydrobiologia 695: 343–360. [CrossRef] [Google Scholar]
- Tudesque L, Chrea S, Chea R, Wetzel CE. 2023. Lucectorea gen. nov., a new atypical genus of biraphid diatoms (Bacillariophyceae) with bent frustule. Nova Hedwigia 17: 25–44. [CrossRef] [Google Scholar]
- Tudesque L, Le Cohu R, Van de VijverF B. 2021. The diatom flora of the Tonle Sap Lake in Cambodia: Centrobacillariophycidae-inventory and description of five new taxa including Praecyclostephanos gen. nov. Diatom Res 36: 185–235. [CrossRef] [Google Scholar]
- Tudesque L, Pool TK, Chevalier M. 2019. Planktonic diatom community dynamics in a tropical flood-pulse lake: the Tonle Sap (Cambodia). Diatom Res 34: 1–22. [CrossRef] [Google Scholar]
- Venkatachalapathy R, Karthikeyan P. 2015. Application of Diatom-Based Indices for Monitoring Environmental Quality of Riverine Ecosystems: A Review BT − Environmental Management of River Basin Ecosystems. (Eds M. Ramkumar, K. Kumaraswamy & R. Mohanraj), pp. 593–619. Cham: Springer International Publishing. [Google Scholar]
- Verleyen E, Vyverman W, Sterken M, Hodgson DA, De Wever A, Juggins S, et al. 2009. The importance of dispersal related and local factors in shaping the taxonomic structure of diatom metacommunities. Oikos 118: 1239–1249. [CrossRef] [Google Scholar]
- Wang Y-K, Stevenson RJ. 2005. Development and evaluation of a diatom-based Index of Biotic Integrity for the Interior Plateau Ecoregion, USA. J North American Benthological Society. 24:, 990–1008. [Google Scholar]
- Warton DI, Wright ST, Wang Y. 2012. Distance-based multivariate analyses confound location and dispersion effects. Methods Ecol Evolut 3: 89–101. [CrossRef] [Google Scholar]
- Whiting K. 2022. Here's how the food and energy crises are connected. Sustainable Development Impact Meetings: The World Economic Forum. 19–23 September 2022, New York, USA. [Google Scholar]
- Wu J. 1999. A generic index of diatom assemblages as bioindicator of pollution in the Keelung River of Taiwan. Hydrobiologia 397: 79–87. [Google Scholar]
- Wu N, Cai Q, Fohrer N. 2012. Development and evaluation of a diatom-based index of biotic integrity (D-IBI) for rivers impacted by run-of-river dams. Ecolog Indic 18: 108–117. [CrossRef] [Google Scholar]
- Yoshikawa T, Takagi AP, Ishikawa S, Hori M, Sitha H, Cheasan E, et al. 2024. Spatial pattern of phytoplankton and its habitat conditions of Tonle Sap Lake in Cambodia during the low-water and high-water seasons. Limnology. https://doi.org/10.1007/s10201-024-00766-5 [Google Scholar]
- Zepernick BN, Gann ER, Martin RM, Pound HL, Krausfeldt LE, Chaffin JD, et al. 2021. Elevated pH conditions associated with Microcystis spp. blooms decrease viability of the cultured diatom fragilaria crotonensis and natural diatoms in lake erie. Front Microbiol 12: 1–12. [CrossRef] [Google Scholar]
Cite this article as: Chrea S, Chea R, Tudesque L. 2024. Spatial patterns of benthic diatom community structure in the largest northwestern river of Cambodia (Sangker River). Knowl. Manag. Aquat. Ecosyst., 425, 21.
All Tables
List of indicator genera of benthic diatom flora by clusters along the Sangker river catchment, Tonle Sap basin (number of permutations = 999).
All Figures
|
Fig. 1 Map of the Sangker River catchment (Tonle Sap Basin, Cambodia), with the geographical location of the nineteen monitoring sites (grey polygones) and the red star indicating the largest city within the catchment area. |
| In the text | |
|
Fig. 2 Distribution by percentages of benthic diatom genera in the Sangker River catchment (Tonle Sap Basin) according to (a) categories, (b) orders, and (c) families. All families with a number of genera less than 3% were grouped in “Others” and shown in the box next to the pie chart. |
| In the text | |
|
Fig. 3 Abundance and occurrence of benthic diatom community along the Sangker River catchment (Tonle Sap Basin). (a) Relative abundance of diatom genera in percentage (only genera with a relative abundance greater than 1% are shown), (b) Occurrence of diatom genera as a percentage (only diatom genera with a frequency of occurrence greater than 50% are shown). Seventy-eight (78) diatom genera were recorded from the nineteen (19) monitoring sites within the river catchment. |
| In the text | |
|
Fig. 4 Spatial distribution of the benthic diatom community along the Sangker River catchment (Tonle Sap Basin). Hierarchical clustering (cluster dendrogram) according to the similarity of diatom genera composition (top left). Polygonal dots represent the diatom monitoring sites associated with the identified cluster (Ia & Ib, II). Different colors indicated benthic diatom community assemblages: red (Ia), yellow (Ib), and blue (II), while the black dot represented the dams. |
| In the text | |
|
Fig. 5 Indicator genera of the diatom community are presented by clusters (Ia, Ib, and II) and by combined clusters (Ia and Ib, Ib and II); only the common species within the indicator genera were selected for illustration. |
| In the text | |
|
Fig. 6 Canonical correspondence analysis (CCA) relating the benthic diatom community (a) to the physicochemical variables (b). Different colors of text on the left plot indicate the indicator genera in each diatom assemblage (yellow text denotes indicator genera from cluster Ia and red and blue from clusters Ib and II, respectively), while the grey texts on the right plot shows the monitoring sites. The blue arrows represent the vectors of physiochemical variables; only significant variables are depicted (Monte Carlo permutation tests with 999 permutations, p-value < 0.05). A detailed description of the benthic diatom genera and Monte Carlo permutation tests are given in Tabs. S1 and S2. |
| In the text | |
|
Fig. 7 Venn diagram of variance partitioning results showing the relative effect of physical and chemical factors alone and in combination with the variation of the benthic diatom community. Numbers represent percentages of variation explained by each factor; all factors were statistically significant (p-value <0.05). |
| In the text | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.