| Issue |
Knowl. Manag. Aquat. Ecosyst.
Number 425, 2024
Development of biological and environmental indicators and indices, testing and use
|
|
|---|---|---|
| Article Number | 1 | |
| Number of page(s) | 12 | |
| DOI | https://doi.org/10.1051/kmae/2023024 | |
| Published online | 23 January 2024 | |
Research Paper
Deep chlorophyll maximum in temperate lakes with different trophic conditions − a rare or common phenomenon?
1
Department of Hydrobiology, Faculty of Biology, University of Białystok, Ciołkowskiego 1J, 15-245 Białystok, Poland
2
Department of Freshwater Protection, Institute of Environmental Protection-National Research Institute, Słowicza 32, 02‑170 Warsaw, Poland
* Corresponding author: This email address is being protected from spambots. You need JavaScript enabled to view it.
Received:
9
September
2023
Accepted:
4
December
2023
Abstract
Deep chlorophyll maxima (DCM) are a common phenomenon in low-trophic stratified lakes, and there is much less information about the occurrence of DCM in eutrophic and dystrophic lakes. Therefore, we performed in situ continuous measurements of chlorophyll concentration by a submersible spectrofluorometer in 23 temperate lakes with different trophic conditions (mesotrophic, eutrophic, and dystrophic). We perform chemical analyses of available nutrient forms to better understand the mechanism of DCM formation. We found the highest concentration of phytoplankton in the metalimnion or upper hypolimnion of most studied lakes, regardless of trophic conditions. Nevertheless, the differences in the number of phytoplankton between DCM and epilimnion were largest in low-trophic lakes, where the chl a concentration in DCM was even 15 times higher than that in the epilimnion. The mesotrophic lakes showed vertical niche segregation by phytoplankton, with a general pattern − green algae in the upper layer, below diatoms, then cryptophytes (and cyanobacteria). Our results indicated that the main factor for DCM in mesotrophic and eutrophic lakes was epilimnetic nutrient depletion, while in dystrophic lakes, it seems to be caused by other factors due to the depletion of dissolved nutrients in whole water profiles.
Key words: Phytoplankton / vertical distribution / trophic conditions / deep chlorophyll maximum / in situ fluorescence
© M. Karpowicz et al., Published by EDP Sciences 2024
 This is an Open Access article distributed under the terms of the Creative Commons Attribution License CC-BY-ND (https://creativecommons.org/licenses/by-nd/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. If you remix, transform, or build upon the material, you may not distribute the modified material.
This is an Open Access article distributed under the terms of the Creative Commons Attribution License CC-BY-ND (https://creativecommons.org/licenses/by-nd/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. If you remix, transform, or build upon the material, you may not distribute the modified material.
1 Introduction
Phytoplankton, the primary producers in aquatic ecosystems, are essential components of lake food webs and play a fundamental role in carbon and nutrient cycling. The vertical distribution of phytoplankton within the water column is influenced by various factors, including light, nutrients, thermal and trophic regimes of the lake, together with zooplankton grazing (Reynolds, 1992). Understanding the vertical distribution patterns of phytoplankton communities is crucial for comprehending lakes’ ecological dynamics and productivity (Gliwicz and Kowalczewski, 1981; Karpowicz and Ejsmont-Karabin, 2017).
In this study, we focused on investigating the occurrence of a specific feature of phytoplankton distribution known as the ‘deep chlorophyll maximum’ (DCM). DCM refers to a phenomenon that consists of the accumulation of photosynthetic microorganisms in a thin layer within the water column (Camacho, 2006). The DCM is often seen in deep lakes of the temperate zone during the summer thermal stratification period when oligotrophic and mesotrophic conditions permit epilimnetic nutrient depletion and offer sufficient light to reach the metalimnion and upper hypolimnion, where nutrient availability is frequently higher (Reynolds, 1992). Therefore, in low trophic conditions, phototrophs need to compromise between being deep enough to have available nutrients and being shallow enough to have enough light. With increasing depth, there is light attenuation caused by water itself, dissolved substances and particular matter, including phytoplankton. As a result, light arriving at deep layers mainly corresponds to the central and upper parts of the photosynthetically active radiation spectrum from 550 to 630 nm (Gervais, 1991; Callieri et al., 1996; Vila et al., 1996). These wavelengths can be harvested by phycoerythrin and phycoerythrocyanin, which favor cyanobacteria and other algae with these photosynthetic pigments to cope with the low light availability in deeper layers (Stockner et al., 2000; Camacho et al., 2001, 2003).
The other factors that could favor the metalimnetic maximum of algae are increased water density and viscosity as a result of the sharp temperature gradient in this layer. This limits sedimentation loss and causes algae to progressively accumulate at a depth where cell density equals water density (neutral buoyancy), which is important especially for non-motile species (Diehl, 2002; Karpowicz and Ejsmont-Karabin, 2017). Therefore, heavy and non-motile diatoms could take advantage of higher water density (Arvola et al., 1991; Reynolds, 1992; Camacho et al., 2001; Wetzel, 2001; Winder et al., 2009). On the other hand, very motile cryptophytes, chrysophytes, and raphidophytes can have mixotrophic feeding strategies (Vincent and Goldman, 1980) and can benefit from higher bacterial biomass and nutrient levels in the upper hypolimnion due to their low light requirements (Ptacnik et al., 2003). Therefore, long-lasting stratification maintains relatively constant conditions in the different water layers creating different habitats. As a consequence, different groups of algae were able to develop intensively at depths where these environmental conditions overlapped with their requirements (Reynolds, 1992; Camacho, 2006).
Our main objectives in this study were twofold. First, we aimed to investigate how thermal stratification influences the vertical distribution of the main taxonomic groups of phytoplankton communities during summer stagnation in lakes of different trophic conditions. The vertical distribution of phytoplankton was determined by in situ and continuous measurements of chlorophyll by a submersible spectrofluorometer. This method provides a great advantage over standard sampling because many species of phytoplankton can achieve the highest concentration in a very thin layer (Gregor and Marsálek, 2004). Lakes with contrasting trophic statuses provide an ideal opportunity to examine the potential influence of nutrient availability on the distribution patterns of phytoplankton along the water column. The factors affecting DCM have been extensively studied in low-trophic lakes, while the occurrence of DCM in eutrophic and dystrophic lakes is less well understood (Gálvez et al., 1988; Pęczuła et al., 2013; Pęczuła et al., 2018; Karpowicz and Ejsmont-Karabin, 2018). Second, we aimed to identify the factors that promote the occurrence of the deep chlorophyll maximum. By examining the relationship between chlorophyll concentrations and environmental factors, such as temperature, light, and dissolved nutrient concentrations, we wanted to determine the drivers that favor the formation and maintenance of the DCM in lakes with different trophic statuses. Our study contributes to a better understanding of the mechanisms governing phytoplankton dynamics in lakes by examining the vertical distribution patterns of phytoplankton and their relationship with environmental factors. This knowledge has implications for the management and conservation of aquatic ecosystems, as well as for predicting the responses of phytoplankton communities to future environmental changes.
2 Study area and methods
The study was conducted in 23 lakes in northeastern Poland (Fig. 1) during the period of stable summer stagnation (second part of July) in 2016–2019 under similar conditions (afternoon 11 a.m. − 3 p.m.; no rainfall). The studies covered lakes with a broad range of maximum depths (3.5–108.5 m) that showed complete or, in a few cases, partial thermal stratification. The studied lakes are characterized by different trophic states; eight of them (including three sites of Lake Wigry) represent mesotrophic conditions (TSI below 50), six lakes are eutrophic (TSI between 50–60), and nine are dystrophic (humic) lakes which are characterized by high HDI values indicating advanced dystrophy (Tab. 1). Furthermore, dystrophic lakes also had other features typical of humic waters: acidity, a small amount of dissolved mineral substances but large dissolved organic carbon (DOC) concentrations, and yellow-brown color (Karpowicz et al., 2023). The dystrophic lakes were small, shallow (max depth ranged from 3.5 to 10 meters), usually oval, without any outlets, and surrounded by forest. The mesotrophic lakes were usually large, with depths ranging from 10 to 108.5 meters, and the visibility of the Secchi disc ranged from 2.4 to 7 meters (Tab. 1). The maximum depth of the eutrophic lakes ranged from 6.4 to 44.5 meters, and the visibility of the Secchi disc ranged from 0.5 to 2.3 meters (Tab. 1).
The sampling stations were located close to the deepest point of each lake, with the exception of Lake Wigry, where we sampled three of its basins separately. The field measurements included Secchi disc visibility (SDV), pH, and electrolytic conductivity (EC) measured using an HQ40D Multi Meter (Hach-Lange GmbH). Temperature and chlorophyll concentrations (total chlorophyll a; green algae, diatoms, cyanobacteria, and cryptophytes) were measured in situ with the submersible spectrofluorometer FluoroProbe (bbe-Moldaenke). Total chlorophyll a constitutes the sum of chlorophyll found in the cells of all phytoplankton groups. Continuous measurements of temperature and phytoplankton every few centimeters allowed us to present a detailed vertical distribution of phytoplankton and precisely determine the thermocline/metalimnion range (temperature change of at least 1 °C per meter or relatively more rapid temperature change) (Wetzel, 2001). FluoroProbe identifies the four phytoplankton groups that represent distinct taxonomic units: green algae (Chlorophyceae and Euglenophyceae), cyanobacteria (phycocyanin-rich Cyanobacteria), diatoms (Heterokontophyta, Haptophyceae, and Dinophyceae), and cryptophytes (Cryptophyta and phycoerythrin-rich Cyanobacteria). A fluorometric evaluation of algal classes is provided by differences in the ensuing chlorophyll emission, which is based on variations in class-dependent peripheral antenna pigments (Beutler et al., 2002; Kring et al., 2014).
The concentrations of dissolved nutrient forms (orthophosphate PO43−, ammonium NH4+, nitrate NO3−, and nitrite NO2−) were determined in the laboratory using Dionex ICS 1100 ion chromatography. Total phosphorus (TP) analyses were conducted according to the standard photocolorimetric method (Neal et al., 2000). The concentrations of dissolved nutrients were analyzed for each thermal layer of lakes. The trophic status of lakes was established using Carlson's trophic state index (TSI) as an average of three parameters: Secchi disc visibility (SDV), chlorophyll a concentration (chl a), and TP (Carlson, 1977). The hydrochemical dystrophy index (HDI) was used to evaluate the state of dystrophy (Górniak, 2017), where values larger than 65 indicate advanced dystrophy. The depth of the euphotic zone was estimated by multiplying SDV by 2.5, the most typical ratio of euphotic zone depth to SDV for lakes in northern Poland (Borowiak and Borowiak, 2016) while for dystrophic lakes a coefficient of 1.5 was used according to Eloranta (1978).
Statistical analyses were performed with XLSTAT Ecology (Addinsoft). Basic descriptive statistics for chlorophyll a and the share of cyanobacteria is presented as box plots (Fig. 2). One-way ANOVA with post-hoc Tukey's HSD (Honest Significant Difference) was used to test all pairwise differences between means. Multiple linear regressions, Spearman correlations, and canonical correspondence analysis (CCA) were used to analyze the effect of dissolved nutrients on different phytoplankton groups.
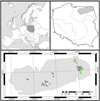 |
Fig. 1 Location of the analyzed lakes in north-eastern Poland. The number of lakes correspond to Table 1, and the color of the dot represents the trophy of the lakes (brown − dystrophic; light green − mesotrophic; dark green − eutrophic). The grey field at the bottom represent East Baltic Lake District in Poland. |
Morphometric and trophic characteristics of the studied lakes in NE Poland. TSI − Carlson trophic state index (Carlson, 1977); HDI − hydrochemistry dystrophy index (Górniak, 2017); SDV − Secchi disk visibility.
 |
Fig. 2 Average chlorophyll a concentrations in lakes with different trophic status (a) and average chl a in water layers (b). Percentage share of cyanobacteria in lakes with different trophic statuses (c) and in water layers (d). The different italics letters (a, b, c) above the box plots indicates significantly different values at p < 0.05, while the same letters indicate no significant differences. The lower and upper limits of the boxes are the first and third quartiles. The red crosses represent the means and the central horizontal bars are the medians. Points above or below are outliers and the whiskers represent minimum and maximum values. |
 |
Fig. 3 Vertical profiles of total chlorophyll a concentrations in mesotrophic lakes with low chl a (a), mesotrophic lakes with higher chl a and Lake Wigry (b), eutrophic lakes (c), and dystrophic lakes (d). |
3 Results
3.1 Temperature, light, and nutrient gradients
We observed differences in the thermal stratification of lakes with different trophic statuses. The strongest stratification was observed in dystrophic lakes, where at a depth of 4 meters the temperature was approximately 10 °C (Fig. S1c). Therefore, the dystrophic lakes had a very shallow epilimnion, and the thermocline began at a depth of 1–2 meters (Fig. 6). Thermocline in shallow eutrophic lakes (no. 9, 11, and 12) started at 3–4 meters, while in deeper eutrophic lakes it started at 5–6 meters (Fig. 5; Fig. 1Sb). The greatest thickness of epilimnion was observed in mesotrophic lakes where a sharp thermal gradient started from 6–7 meters (Fig. 1Sa), except for lakes no. 5 and no.6, where the thermal gradient began from 3.5 and 5 meters, respectively (Fig. 4).
Despite differences in trophic conditions, most lakes were characterized by a very good light regime in the metalimnion zone; moreover, in some cases, light also penetrated the upper layers of hypolimnion. Among the mesotrophic lakes, worse light conditions were found only in lake no. 3 (Fig. 4). In eutrophic lakes, SDV measurements indicated a shallow extent of the euphotic zone (limited within the epilimnion layer) in four lakes: 10, 11, 13, and 14 (Fig. 5). The dystrophic lakes were characterized by the extent of the light zone reaching the depths within the metalimnion (Fig. 6).
Dystrophic lakes in all layers had a low concentration of dissolved forms of nitrogen and phosphorus, except for higher concentrations of ammonium ions in the hypolimnion (Tab. 2). Eutrophic lakes were characterized by a low concentration of dissolved forms of nutrients in the epilimnion zones, and much higher concentrations of PO43−, NH4+, and NO3− in the metalimnion and hypolimnion zones (Tab. 2). Mesotrophic lakes had similar concentrations of NH4+ and NO2− in the vertical profiles, and higher concentrations of orthophosphates in the metalimnion zones, as well as higher concentrations of nitrate in the metalimnion and hypolimnion zones (Tab. 2).
 |
Fig. 4 Vertical distribution of green algae, cyanobacteria, diatoms, and cryptophytes in mesotrophic lakes. The gray fields represent the metalimnion boundaries and the dotted lines are the borders of the euphotic zones. In the upper left corner, the numbers of lakes (no.) correspond to Table 1. |
Vertical variation of dissolved nutrient concentrations in lakes with different trophic status.
3.2 Total chlorophyll a concentrations and share of cyanobacteria
There were statistically significant differences in total chlorophyll a concentrations in lakes of different trophic statuses (F = 19.1; p < 0.001). The average concentrations of chl a in mesotrophic, eutrophic, and dystrophic lakes were 5.6 ± 4.1 μg L−1, 21.2 ± 25.0 μg L−1, and 38.3 ± 29.5 μg L−1, respectively (Fig. 2a). The highest concentrations of chl a in the vertical profiles were found in the metalimnion, and there were no statistically significant differences in chl a concentrations in the epilimnion and hypolimnion (Fig. 2b). The lowest share of cyanobacteria was observed in dystrophic lakes, and there were no statistically significant differences in the share of cyanobacteria between eutrophic and mesotrophic lakes (Fig. 2c). The share of cyanobacteria increased with depth and the highest share was found in the hypolimnion (Fig. 2d).
The highest chlorophyll concentrations were observed mostly in the metalimnion or upper hypolimnion despite differences in trophic conditions of lakes (Figs. 3–6). In the lake with the lowest trophic level (no. 1), the maximum chlorophyll concentration was at 11 meters with 64.7 μg L−1 (approximately 15 times higher than in the epilimnion), in lake no. 7c the maximum was at 10 meters with 33.4 μg L−1 (approximately 5 times higher than in the epilimnion), in lake no. 8 the maximum was at 11 meters with 17.6 μg L−1 (approximately 5 times higher than in the epilimnion) (Fig. 3b); and in lake no. 2, the maximum was at 11 meters with 9.0 μg L−1 (approximately 3 times higher than in the epilimnion) (Fig. 3a). The eutrophic lakes also had a maximum concentration of phytoplankton in the metalimnion zones (Figs. 3c and 5), except lake no. 10 which had maximum concentrations of algae at 4 meters (Fig. 5). All dystrophic lakes had a phytoplankton maximum in the metalimnion (Figs. 3d and 6).
 |
Fig. 5 Vertical distribution of green algae, cyanobacteria, diatoms, and cryptophytes in eutrophic lakes. The gray fields represent the metalimnion boundaries and the dotted lines are the borders of the euphotic zones. In the upper left corner, the numbers of lakes (no.) correspond to Table 1. |
 |
Fig. 6 Vertical distribution of green algae, cyanobacteria, diatoms, and cryptophytes in dystrophic lakes. The gray fields represent the metalimnion boundaries and the dotted lines are the borders of the euphotic zones. In the upper left corner, the numbers of lakes (no.) correspond to Table 1. |
3.3 Vertical niche segregation by the main group of phytoplankton
The mesotrophic lakes exhibited distinctive vertical niche segregation among phytoplankton, where green algae prevailed in the upper layer, while diatoms, cryptophytes, and cyanobacteria dominated the lower layers (Fig. 4). The deep chlorophyll maxima in mesotrophic lakes were caused by cryptophytes and diatoms (lakes no. 2, 6, 7a, 7b, 7c, 8) or cryptophytes and cyanobacteria (lakes no. 1 and no. 3) (Fig. 4).
The deep chlorophyll maxima in eutrophic lakes were caused by different groups of algae (Fig. 5). The diatoms created DCM in lakes no. 10 and 13; while cyanobacteria and cryptophytes created DCM in lakes no. 11 and 14 (Fig. 5). There was no clear vertical niche segregation by phytoplankton in eutrophic lakes (Fig. 5). Dystrophic lakes were dominated by green algae, with the highest concentrations in the metalimnion zones (Fig. 6). In the metalimnion, in most of the studied lakes, the share of algae from the diatoms group increased, which also includes algae from the Heterokontophyta phylum (including Raphidophyceae common in dystrophic lakes).
3.4 Environmental factors affect the vertical distribution of phytoplankton
The limitation of dissolved forms of nutrients in the epilimnion of lakes in different trophic conditions seems to be the main factor influencing the occurrence of DCM (Tab. 2). The total chlorophyll a concentrations were negatively correlated with PO43− (−0.515; p <0.0001), NO2− (−0.442; p = 0.0002), and NO3− (−0.354; p = 0.003). Moreover, in mesotrophic and dystrophic lakes, nutrient limitation was observed throughout the entire vertical profile. Only eutrophic lakes were characterized by high concentrations of orthophosphates and nitrates in the metalimnion and hypolimnion zones (Tab. 2). The CCA analysis in eutrophic and mesotrophic lakes suggests that cryptophytes are related to PO43− and NO2−, while cyanobacteria seem to be related to NO3− (Fig. 7a). The green algae and diatoms in eutrophic and mesotrophic lakes seem to be related to temperature, DOC, and NH4+ (Fig. 7a).
The dystrophic lakes with the highest concentrations of chlorophyll a (Fig. 2a) were characterized by very low concentrations of dissolved forms of nutrients in the water column (Tab. 2). Nevertheless, in all dystrophic lakes, a metalimnetic maximum of phytoplankton was observed (Fig. 6). The CCA analysis confirms that in dystrophic lakes dominant green algae were weakly related to dissolved nutrient concentration and other environmental factors (temperature, pH, DOC, EC) (Fig. 7b). In dystrophic lakes, diatoms could be related to PO43−, NH4+, DOC, and EC, while cyanobacteria and cryptophytes seem to be related to NO2− and pH (Fig. 7b).
 |
Fig. 7 Canonical correspondence analysis map of a major group of phytoplankton and nutrient forms and environmental variables in eutrophic and mesotrophic lakes (a); and dystrophic lakes (b). |
4 Discussion
The results of our study pointed out the highest concentration of phytoplankton in the metalimnion or upper hypolimnion of most studied lakes with different trophic statuses (mesotrophic, eutrophic, dystrophic). A deep chlorophyll maximum (or deep chlorophyll layer) is a common phenomenon in oligotrophic and mesotrophic temperate lakes (e.g., Fee, 1976; Barbiero and Tuchman, 2004; Camacho, 2006; Scofield et al., 2020; Pasztaleniec and Ochocka, 2021), as well as in the global ocean and seas (e.g., Ediger and Yilmaz, 1996; Cullen et al., 2015; Cornec et al., 2020). The results of our study showed the occurrence of DCM in eutrophic and dystrophic lakes, which has rarely been reported (Gálvez et al., 1988; Karpowicz and Ejsmont-Karabin, 2018; Caballero and Vázquez, 2020). Nevertheless, in low-trophic lakes, we observed that the differences in chlorophyll concentrations between DCM and epilimnion were much higher, with DCM exhibiting chlorophyll concentrations up to 15 times higher than those of epilimnion.
The maximum density of algae, expressed by the concentration of chlorophyll a was formed within the metalimnion, or slightly below, and its location usually coincided with the maximum range of the euphotic zone. The phytoplankton maximum often occurs at the depth where light and nutrient availability are equal, according to models of the vertical distribution of phytoplankton in stratified mixed water columns (Klausmeier and Litchman, 2001). These models also emphasized that a key factor influencing phytoplankton dynamics is the light:nutrient ratio (Huisman and Weissing, 1995; Diehl, 2002). Our results confirmed that dissolved nutrient limitation in epilimnion zones could be the main factor responsible for DCM in mesotrophic and eutrophic lakes, where metalimnion and hypolimnion were nutrient-rich layers (Tab. 2). However, the metalimnetic phytoplankton maximum in dystrophic lakes seems to be caused by other factors, because their metalimnion was a nutrient-poor layer (Tab. 2). Mixotrophic and osmotrophic phytoplankton are frequently the main constituents of primary producers in dystrophic lakes, and they may benefit from increased bacterial biomass and DOC concentrations (Johansson et al., 2013; Gollnisch et al., 2021; Karpowicz et al., 2023; Rohrlack, 2023).
Some phytoplankton species can also migrate downward to avoid bright illumination on sunny days. Many flagellate algae migrate diurnally within the epilimnion or even between the epilimnion and hypolimnion (Camacho, 2006). Common species found in humic waters frequently employ this movement strategy, including Cryptomonas ovata and Cryptomonas marssonii (Arvola, 1984; Smolander and Arvola, 1988), Gonyostomum semen (Pęczuła et al., 2013, 2018), and Chlamydomonas spp. (Jones, 1988). The Gonyostomum semen movement ability is estimated at 1 m/h, and this species is capable of overcoming a temperature gradient of 10 °C (Jones, 1988; Pęczuła et al., 2013). The presence of G. semen as well as an increase in its abundance were observed in most studied lakes (Pęczuła et al., 2018; Karpowicz et al., 2023). Our previous research on dystrophic lakes using both submersible spectrofluorometer FluoroProbe and taxonomic recognition indicates that G. semen was probably classified by a probe as a green algae (Karpowicz et al., 2020; Karpowicz et al., 2023).
Our research also showed that maximum phytoplankton densities occurred most often near the edge of the photic zone regardless of trophic conditions. Therefore, the nutrient-rich metalimnion with sufficient light and higher water density seems to be an ideal place for the development of phytoplankton during summer stagnation. This layer is also an important food source for zooplankton, which contributes to the efficient transfer of matter and energy in planktonic food webs (Karpowicz et al., 2021). Therefore, the presence of DCM is the preferred phenomenon during summer stagnation, which points to the limitation of dissolved nutrients in the epilimnion and thus indicates a good ecological status of lakes. On the other hand, some research suggests that DCM of clean lakes in meta- and hypolimnion may lead to misclassification of the lake ecological status by one or even two status classes in a 5-class assessment system (Nõges et al., 2010; Pasztaleniec and Ochocka, 2021). Our research suggests that this approach may strongly overestimate the ecological status of clean lakes due to the presence of DCM. Therefore, our data confirm that current indicators of ecological status based on phytoplankton properly consider only the epilimnion or photic zone (Poikane, 2009; Wolfram and Dokulil, 2009).
In mesotrophic lakes, we found vertical niche segregation by phytoplankton, with a general pattern − green algae in the upper layer, below diatoms, then cryptophytes (and cyanobacteria). The green algae reached the highest density in the epilimnion due to their high light requirements (Litchman, 2000). Diatoms and cryptophytes reached higher densities in nutrient-rich lower water layers, with maximum densities noted at different depths. Fast-sinking diatoms (heavy and non-motile) could also benefit from greater water density (Arvola et al., 1991; Reynolds, 1992; Winder et al., 2009), as well as from higher concentrations of silica in lower water layers (Gibson et al., 2000; Shatwell et al., 2013). Dinophytes could also be a group of algae forming the recorded chlorophyll maxima of the studied meso- and eutrophic lakes because they have similar to diatom pigments emission spectra measured by spectrofluorometer. Many studies have shown that some dinophytes occur preferentially in deep layers of lakes with low nutrient concentrations which is facilitated by their motility, mixotrophy, and physiological adaptation to low light (e.g., Grigorszky et al., 2003; Solis et al., 2013). Cryptophytes are also very motile and can have mixotrophic feeding strategies (Vincent and Goldman, 1980), and due to their low light requirements, they can benefit from greater nutrient levels and bacterial biomass found at the edge of the hypolimnion (Ptacnik et al., 2003). Furthermore, the presence of phycobilins gives cyanobacteria and cryptophytes a selective advantage to deal with the limited light availability at deep layers, which mostly correspond to the spectrum from 550 to 630 nm (Gervais, 1991; Callieri et al., 1996; Vila et al., 1996). For many cyanobacterial taxa, the ability to change their cell buoyancy is another factor that facilitates movement. Together with the above mentioned photoadaptation abilities, these factors are responsible for the common occurrence of cyanobacteria in the metalimnetic layer of mesotrophic lakes as was also observed in Polish lakes (Wojciechowska et al., 2004; Solis et al., 2013; Pasztaleniec and Ochocka, 2021; Lenard and Poniewozik, 2022). On the other hand, in most eutrophic lakes, there was no clear vertical niche segregation by phytoplankton, and most algae reached higher density in the metalimnion, thereby creating a DCM. This may be a result of dissolved nutrient depletion in the epilimnion, which was also reported from eutrophic lakes without external anthropogenic discharges as a result of internal nutrient cycling (Wu et al., 2017; Wang et al., 2019).
Our results provide valuable insights into the distribution patterns of phytoplankton in lakes with different trophic conditions and shed light on the potential factors influencing the formation of DCM. Furthermore, our results confirm that in situ and continuous measurements of chlorophyll are excellent tools for studying the formation of DCM, and provide a great advantage over standard sampling because many species of phytoplankton can achieve the highest concentration in a very thin layer (Gregor and Marsálek, 2004). We pointed out that the submersible spectrofluorometer is an efficient and easy-to-use tool in the real-time monitoring of phytoplankton communities and could act as an early warning system for the detection of potentially harmful algae blooms (Garrido et al., 2019). The usefulness of real-time measurements, giving quick insight into phytoplankton, as an alarm system or in the event of ecological disasters, was also pointed out by other authors (Kring et al., 2014; Garrido et al., 2019; Pasztaleniec et al., 2020). It seems that it could also be a great tool for evaluating ecological status because it shows the total chlorophyll a concentration and the percentage of cyanobacteria, and these parameters are crucial when assessing the ecological status of lakes (Hutorowicz and Pasztaleniec, 2021). Unlike traditional monitoring, which is time-consuming (sampling and long time required for microscopic analysis) and requires extensive taxonomic knowledge to correctly determine the composition of phytoplankton.
Competing interests
The authors have no competing interests to declare that are relevant to the content of this article.
Data availability statement
All data generated or analyzed during this study are included in this published article and its supplementary information file.
Author contributions
M. Karpowicz wrote the main manuscript text, performed statistical analyses, prepared figures, conceptualization, and investigation; Pasztaleniec A. writing − review & editing; Grabowska M. writing − review & editing, investigation; Pietryczuk A. investigation. All authors reviewed the manuscript.
Supplementary Material
Figure S1. Vertical profiles of temperature in mesotrophic trophic lakes (a), eutrophic lakes (b), and dystrophic lakes (c). Access Supplementary Material
Acknowledgments
The authors are grateful to Adam Więcko for dissolved nutrient analyses and Sebastian Kutyła for the preparation map of the study site. The authors are also grateful to the editor and reviewers for their time and energy in providing helpful comments that have improved this manuscript. This article has received financial support from the Polish Ministry of Science and Higher Education under a subsidy for maintaining the research potential of the Faculty of Biology, University of Bialystok.
References
- Arvola L. 1984. Diel variation in primary production and the vertical distribution of phytoplankton in a polyhumic lake. Arch Hydrobiol 101: 503–519. [Google Scholar]
- Arvola L, Ojala A, Barbosa F, Heaney SI. 1991. Migration behavior of three cryptophytes in relation to environmental gradients: an experimental approach. Brit Phycol J 26: 361–373. [CrossRef] [Google Scholar]
- Barbiero RP, Tuchman ML. 2004. The deep chlorophyll maximum in Lake Superior. J Great Lakes Res 30: 256–268. [CrossRef] [Google Scholar]
- Beutler M, Wiltshire KH, Meyer B, Moldaenke C, Lüring C, Meyerhöfer M, Hansen U-P, Dau H. 2002. A fluorometric method for the differentiation of algal populations in vivo and in situ. Photosynth Res 72: 39–53. [CrossRef] [PubMed] [Google Scholar]
- Borowiak D, Borowiak M. 2016. Comparative studies of underwater light regimes in lakes of the East-Suwałki Lakeland. Limn Rev 16: 173–183. [Google Scholar]
- Caballero M, Vázquez G. 2020. Mixing patterns and deep chlorophyll a maxima in a eutrophic tropical lake in western Mexico. Hydrobiologia 847: 4161–4176. [CrossRef] [Google Scholar]
- Callieri C, Amicucci E, Bertoni R, Vörös L. 1996. Fluorometric characterization of two picocyanobacteria strains from lakes of different underwater light quality. Int Revue ges Hydrobiol 81: 13–23. [CrossRef] [Google Scholar]
- Camacho A. 2006. On the occurrence and ecological features of deep chlorophyll maxima (DCM) in Spanish stratified lakes. Limnetica 25: 453–478. [CrossRef] [Google Scholar]
- Camacho A, Vicente E, Miracle MR. 2001. Ecology of Cryptomonas at the chemocline of a karstic sulphate-rich lake. Mar Freshwater Res 52: 805–815. [CrossRef] [Google Scholar]
- Camacho A, Miracle MR, Vicente E. 2003. Which factors determine the abundance and distribution of picocyanobacteria in inland waters? A comparison among different types of lakes and ponds. Arch Hydrobiol 157: 321–338. [CrossRef] [Google Scholar]
- Carlson RE. 1977. A trophic state index for lakes. Limnol Oceanogr 22: 361–369. [Google Scholar]
- Cornec M, Claustre H, Mignot A, Guidi L, Lacour L, Poteau A, D'Ortenzio F, Gentili B, Schmechtig C. 2020. Deep chlorophyll maxima in the global ocean: occurrences, drivers and characteristics. Global Biogeochem Cycles 35: e2020G B006759. [Google Scholar]
- Cullen JJ. 2015. Subsurface chlorophyll maximum layers: Enduring enigma or mystery solved? Annu Rev Mar Sci 7: 207–239. [CrossRef] [PubMed] [Google Scholar]
- Diehl S. 2002. Phytoplankton, light, and nutrients in a gradient of mixing depths: Theory. Ecology 83: 386–398. [CrossRef] [Google Scholar]
- Ediger D, Yilmaz A. 1996. Characteristics of deep chlorophyll maxima in the Northeastern Mediterranean with respect to environmental conditions. J Mar Biol 9: 291–303. [Google Scholar]
- Eloranta P. 1978. Light penetration in different types of lakes in Central Finland. Holarct Ecol 1: 362–366. [Google Scholar]
- Fee EJ. 1976. The vertical and seasonal distribution of chlorophyll in lakes of the Experimental Lakes Area, northwestern Ontario: Implications for primary production estimates. Limnol Oceanogr 21: 767–783. [CrossRef] [Google Scholar]
- Gálvez JA, Niell FX, Lucena J. 1988. Description and mechanism of formation of a deep chlorophyll maximum due to Ceratium hirundinella (O. F. Müller). Arch Hydrobiol 112: 143–155. [CrossRef] [Google Scholar]
- Garrido M, Cecchi P, Malet N, Bec C, Torre F, Pasqualini V. 2019. Evaluation of FluoroProbe® performance for the phytoplankton-based assessment of the ecological status of Mediterranean coastal lagoons. Environ Monit Assess 191: 204. [CrossRef] [MathSciNet] [PubMed] [Google Scholar]
- Gervais F. 1991. Which factors control seasonal and spatial distribution of phytoplankton species in Schlachtensee (Berlin, FRG.)? Arch Hydrobiol 121: 43–65. [CrossRef] [Google Scholar]
- Gibson CE, Wang G, Foy RH. 2000. Silica and diatom growth in Lough Neagh: The importance of internal recycling. Freshw Biol 45: 285–293. [CrossRef] [Google Scholar]
- Gliwicz ZM, Kowalczewski A. 1981. Epilimnetic and hypolimnetic symptoms of eutrophication in Great Mazurian Lakes, Poland. Freshw Biol 11: 425–435. [CrossRef] [Google Scholar]
- Gollnisch RT, Alling T, Stockenreiter M, Ahrén D, Grabowska M, Rengefors K. 2021. Calcium and pH interaction limits bloom formation and expansion of a nuisance microalga. Limnol Oceanogr 66: 3523–3534. [CrossRef] [Google Scholar]
- Górniak A. 2017. A new version of the Hydrochemical Dystrophy Index to evaluate dystrophy in lakes. Ecol Indic 78: 566–573. [CrossRef] [Google Scholar]
- Gregor J, Marsálek B. 2004. Freshwater phytoplankton quantification by chlorophyll a: a comparative study of in vitro, in vivo and in situ methods. Water Res 38: 517–522. [CrossRef] [PubMed] [Google Scholar]
- Grigorszky I, Padisák J, Borics G, Schitchen C, Borbély G. 2003. Deep chlorophyll maximum by Ceratium hirundinella (O. F. Müller) Bergh in a shallow oxbow in Hungary. Hydrobiologia 506: 209–212. [CrossRef] [Google Scholar]
- Huisman J, Weissing FJ. 1995. Competition for nutrients and light in a mixed water column— a theoretical analysis. Am Nat 146 : 536–564. [CrossRef] [Google Scholar]
- Hutorowicz A, Pasztaleniec A. 2021. Uncertainty in phytoplankton-based lake ecological status classification: implications of sampling frequency and metric simplification. Ecol Indic 127: 107754. [CrossRef] [Google Scholar]
- Johansson KSL, Vrede T, Lebret K, Johnson RK. 2013. Zooplankton feeding on the nuisance flagellate Gonyostomum semen. PLoS ONE 8: e 62557. [Google Scholar]
- Jones RI. 1988. Vertical distribution and diel migration of flagellated phytoplankton in a small humic lake. Hydrobiologia 161: 75–87. [CrossRef] [Google Scholar]
- Karpowicz M, Ejsmont-Karabin J. 2017. Effect of metalimnetic gradient on phytoplankton and zooplankton (Rotifera, Crustacea) communities in different trophic conditions. Environ Monit Assess 189: 367. [CrossRef] [PubMed] [Google Scholar]
- Karpowicz M, Ejsmont-Karabin J. 2018. Influence of environmental factors on vertical distribution of zooplankton communities in humic lakes. Ann Limnol − Int J Lim 54: 17. [CrossRef] [EDP Sciences] [Google Scholar]
- Karpowicz M, Zieliński P, Grabowska M, Ejsmont-Karabin J, Kozłowska J, Feniova I. 2020. Effect of eutrophication and humification on nutrient cycles and transfer efficiency of matter in the freshwater food web. Hydrobiologia 847: 2521–2540. [CrossRef] [Google Scholar]
- Karpowicz M, Feniova I, Gladyshev MI, Ejsmont-Karabin J, Górniak A, Sushchik NN, Anishchenko OV, Dzialowski AR. 2021. Transfer efficiency of carbon, nutrients, and polyunsaturated fatty acids in planktonic food webs under different environmental conditions. Ecol Evol 11: 8201–8214. [CrossRef] [PubMed] [Google Scholar]
- Karpowicz M, Grabowska M, Ejsmont-Karabin J, Ochocka A. 2023. Humic lakes with inefficient and efficient transfer of matter in planktonic food webs. Sci Rep 13: 7913. [CrossRef] [PubMed] [Google Scholar]
- Klausmeier ChA, Litchman E. 2001. Algal games: The vertical distribution of phytoplankton in poorly mixed water columns. Limnol Oceanogr 46: 1998–2007. [CrossRef] [Google Scholar]
- Kring SA, Figary SE, Boyer GL, Watson SB, Twiss MR. 2014. Rapid in situ measures of phytoplankton communities using the bbe FluoroProbe: evaluation of spectral calibration, instrument intercompatibility, and performance range. Can J Fish Aquat Sci 71: 1087–1095. [CrossRef] [Google Scholar]
- Lenard T, Poniewozik M. 2022. Planktothrix agardhii versus Planktothrix rubescens: separation of ecological niches and consequences of cyanobacterial dominance in freshwater. Int J Environ Res Public Health 19: 14897. [CrossRef] [PubMed] [Google Scholar]
- Litchman E. 2000. Growth rates of phytoplankton under fluctuating light. Freshw Biol 44: 223–235. [CrossRef] [Google Scholar]
- Neal C, Neal M, Wickham H. 2000. Phosphate measurement in natural waters: two examples of analytical problems associated with silica interference using phosphomolybdic acid methodologies. Sci Total Environ 251: 513–542. [Google Scholar]
- Nõges P, Poikane S, Kõiv T, Nõges T. 2010. Effect of chlorophyll sampling design on water quality assessment in thermally stratified lakes. Hydrobiologia 649: 157–170. [CrossRef] [Google Scholar]
- Pasztaleniec A, Hutorowicz A, Napiórkowska-Krzebietke A. 2020. Rapid monitoring of cyanobacteria in lakes − a case study in the Wel River catchment, Poland. Limnol Rev 20: 41–49. [CrossRef] [Google Scholar]
- Pasztaleniec A, Ochocka A. 2021. A comparative study of phytoplankton epi- and metalimnetic communities under different light and thermal regimes. Ecohydrol Hydrobiol 21: 760–774. [CrossRef] [Google Scholar]
- Pęczuła W, Mencfel R, Kowalczyk-Pecka D. 2013. Among-lake variation in vertical distribution of invasive, bloom-forming algal species Gonyostomum semen (Raphidophyceae) in stratified humic lakes of eastern Poland. Int Rev Hydrobiol 99: 317–325. [Google Scholar]
- Pęczuła W, Grabowska M, Zieliński P, Karpowicz M, Danilczyk M. 2018. Vertical distribution of expansive, bloom-forming algae Gonyostomum semen vs. plankton community and water chemistry in four small humic lakes. Knowl Manag Aquat Ecosyst 419: 28. [Google Scholar]
- Poikane S. 2009. Water Framework Directive intercalibration technical report. Part 2: Lakes. EUR 28838 EN/2, Office for Official Publications of the European Communities, Luxembourg. https://data.europa.eu/doi/10.2788/23415 [Google Scholar]
- Ptacnik R, Diehl S, Berger S. 2003. Performance of sinking and nonsinking phytoplankton taxa in a gradient of mixing depths. Limnol Oceanogr 48: 1903–1912. [CrossRef] [Google Scholar]
- Reynolds CS. 1992. Dynamics, selection and composition of phytoplankton in relation to vertical structure in lakes. Arch Hydrobiol Beih Ergebn Limnol 35: 13–31. [Google Scholar]
- Rohrlack T. 2023. Can osmotrophy in Gonyostomum semen explain why lake browning drives an expansion of the alga in parts of Europe? Limnologica 101: 126097. [CrossRef] [Google Scholar]
- Shatwell T, Kohler J, Nicklisch A. 2013. Temperature and photoperiod interactions with silicon-limited growth and competition of two diatoms. J Plankton Res 35: 957–971. [CrossRef] [Google Scholar]
- Smolander U, Arvola L. 1988. Seasonal variation in the diel vertical distribution of the migratory alga Cryptomonas marssonii (Cryptophyceae) in a small, highly humic lake. In Jones RI, Ilmavirta V. eds. Flagellates in Freshwater Ecosystems. Developments in Hydrobiology Vol. 45, Dordrecht: Springer. https://doi. org/10. 1007/978-94-009-3097-1_8 [Google Scholar]
- Stockner J, Callieri C, Cronberg G. 2000. Picoplankton and other non-bloom-forming cyanobacteria in lakes. In Whitton BA, Potts M. eds. The Ecology of Cyanobacteria. Dordrecht: Springer. https://doi. org/10. 1007/0-306-46855-7_7 [Google Scholar]
- Scofield AE, Watkins JM, Osantowski E, Rudstam LG. 2020. Deep chlorophyll maxima across a trophic state gradient: A case study in the Laurentian Great Lakes. Limnol Oceanogr 65: 2460–2484. [CrossRef] [PubMed] [Google Scholar]
- Solis M, Wojciechowska W, Lenard T. 2013. Vertical distribution of phytoplankton in two mesotrophic lakes. Annales UMSC Sec. C, Vol. LXVIII, 2. https://doi.org/10.2478/v10067-012- 0035-7. [Google Scholar]
- Vila X, Dokulil M, García-Gil LJ, Abellà CA, Borrego CM, Bañeras L. 1996. Composition and distribution of phototrophic bacterioplankton in the deep communities of several central European lakes: the role of light quality. Arch Hydrobiol Beih Ergebn Limnol 48: 183–196. [Google Scholar]
- Vincent WF, Goldman CR. 1980. Evidence for algal heterotrophy in Lake Tahoe, California-Nevada. Limnol Oceanogr 25: 89–99. [CrossRef] [Google Scholar]
- Wang M, Xu X, Wu Z, Zhang X, Sun P, Wen Y, Wang Z, Lu X, Zhang W, Wang X, Tong Y. 2019. Seasonal pattern of nutrient limitation in a eutrophic lake and quantitative analysis of the impacts from internal nutrient cycling. Environ Sci Technol 53: 13675–13686. [CrossRef] [PubMed] [Google Scholar]
- Wetzel RG. 2001. Limnology. San Diego, CA: Academic Press, pp.1006. [Google Scholar]
- Winder M, Reuter JE, Schladow SG. 2009. Lake warming favours small-sized planktonic diatom species. Proc R Soc B: Biol Sci 276: 427–435. [CrossRef] [PubMed] [Google Scholar]
- Wojciechowska W, Poniewozik M, Pasztaleniec A. 2004. Vertical distribution of dominant cyanobacteria species in three lakes − Evidence of tolerance to different turbulence and oxygen conditions. Pol J Ecol 52: 347–351. [Google Scholar]
- Wolfram G, Dokulil MT. 2009. Leitfaden zur Erhebung der Biologischen Qualitätselemente, Seen. Teil B2-01d −Phytoplankton. Handbuch des BMLFUW & des BAW, Wien: pp. 48 . [Google Scholar]
- Wu Z, Liu Y, Liang Z, Wu S, Guo H. 2017. Internal cycling, not external loading, decides the nutrient limitation in eutrophic lake: A dynamic model with temporal Bayesian hierarchical inference. Water Res 116: 231–240. [CrossRef] [PubMed] [Google Scholar]
Cite this article as: Karpowicz M, Pasztaleniec A, Grabowska M, Pietryczuk A. 2023. Deep chlorophyll maximum in temperate lakes with different trophic conditions − a rare or common phenomenon? Knowl. Manag. Aquat. Ecosyst., 425, 1
All Tables
Morphometric and trophic characteristics of the studied lakes in NE Poland. TSI − Carlson trophic state index (Carlson, 1977); HDI − hydrochemistry dystrophy index (Górniak, 2017); SDV − Secchi disk visibility.
Vertical variation of dissolved nutrient concentrations in lakes with different trophic status.
All Figures
|
Fig. 1 Location of the analyzed lakes in north-eastern Poland. The number of lakes correspond to Table 1, and the color of the dot represents the trophy of the lakes (brown − dystrophic; light green − mesotrophic; dark green − eutrophic). The grey field at the bottom represent East Baltic Lake District in Poland. |
| In the text | |
|
Fig. 2 Average chlorophyll a concentrations in lakes with different trophic status (a) and average chl a in water layers (b). Percentage share of cyanobacteria in lakes with different trophic statuses (c) and in water layers (d). The different italics letters (a, b, c) above the box plots indicates significantly different values at p < 0.05, while the same letters indicate no significant differences. The lower and upper limits of the boxes are the first and third quartiles. The red crosses represent the means and the central horizontal bars are the medians. Points above or below are outliers and the whiskers represent minimum and maximum values. |
| In the text | |
|
Fig. 3 Vertical profiles of total chlorophyll a concentrations in mesotrophic lakes with low chl a (a), mesotrophic lakes with higher chl a and Lake Wigry (b), eutrophic lakes (c), and dystrophic lakes (d). |
| In the text | |
|
Fig. 4 Vertical distribution of green algae, cyanobacteria, diatoms, and cryptophytes in mesotrophic lakes. The gray fields represent the metalimnion boundaries and the dotted lines are the borders of the euphotic zones. In the upper left corner, the numbers of lakes (no.) correspond to Table 1. |
| In the text | |
|
Fig. 5 Vertical distribution of green algae, cyanobacteria, diatoms, and cryptophytes in eutrophic lakes. The gray fields represent the metalimnion boundaries and the dotted lines are the borders of the euphotic zones. In the upper left corner, the numbers of lakes (no.) correspond to Table 1. |
| In the text | |
|
Fig. 6 Vertical distribution of green algae, cyanobacteria, diatoms, and cryptophytes in dystrophic lakes. The gray fields represent the metalimnion boundaries and the dotted lines are the borders of the euphotic zones. In the upper left corner, the numbers of lakes (no.) correspond to Table 1. |
| In the text | |
|
Fig. 7 Canonical correspondence analysis map of a major group of phytoplankton and nutrient forms and environmental variables in eutrophic and mesotrophic lakes (a); and dystrophic lakes (b). |
| In the text | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.